Article Text

Statistics from Altmetric.com
The hypothalamus, pituitary, and adrenal glands are dynamic endocrine organs during fetal development.1-3 The adrenal glands, in particular, exhibit remarkable transformation in size, morphology, and function during the prenatal and neonatal periods.2 ,3 It is now recognised that normal development of the hypothalamic–pituitary–adrenal (HPA) axis is essential for: (1) the regulation of intrauterine homeostasis; and (2) the timely differentiation and maturation of vital organ systems including the lungs, liver, and central nervous system necessary for immediate neonatal survival after birth. In addition, acting together with the placenta, the HPA axis might indirectly control the normal timing of parturition in primates.1-3 The liberal use of exogenous antenatal and postnatal corticosteroids during pregnancy and early neonatal life have also raised concerns about potential adverse effects on the HPA axis and subsequent neurodevelopment.4 Thus, an understanding of the physiology and function of the HPA axis in intrauterine and extrauterine life is important for neonatologists. This article aims to provide an overview on the physiology of the glucocorticoid axis and the effects that exogenous corticosteroids have on this system.
Basic physiology
Hormone activity in the HPA axis can be detected between eight and 12 weeks of gestation, early in fetal development.1-3Corticotrophin releasing hormone (CRH) is produced from the fetal hypothalamus and the placenta during pregnancy. It is the primary secretagogue controlling pro-opiomelanocortin (POMC) mRNA expression and pituitary corticotroph secretion of adrenocorticotrophin (ACTH).1 ,3 CRH regulates the growth of pituitary corticotrophs, adrenocortical differentiation, and steroidogenic maturation of the fetal HPA axis.3 It is also a potent vasodilator of the fetoplacental circulation and can potentiate the function of local mediators and hormones, such as prostaglandins and oxytocin, in increasing myometrial contractility during labour.1 The progressive increase in the concentration of CRH in fetal and maternal circulations at late gestation suggests a pivotal role of placental CRH in modulating the timing of parturition (see below).3 ACTH is principally produced by the anterior pituitary corticotrophs and is the prime trophic hormone controlling fetal adrenocortical growth, differentiation, and steriodogenesis.1 ,3 ACTH acts via local mediators or growth factors, such as the vascular endothelial growth factor and epidermal growth factor, in synchronising fetal adrenocortical growth and angiogenesis.
The functional development of the fetal adrenal cortex is a highly complex process that involves the ontogenical expression of specific steroidogenic enzymes in different zones and at different times in gestation.3 The fetal zone is the principal site of dehydroepiandrosterone sulphate (DHEA-S) production, whereas the definitive zone is the main site of mineralocorticoid synthesis. The transitional zone is functionally similar to the fetal zone early in development and is believed to be the site of de novo cortisol production after 28 weeks of gestation.3 Cortisol is important in maintaining intrauterine homeostasis. It also influences the structural and functional development of a wide variety of fetal tissues, and is essential for the antepartum maturation of organ systems including the lungs, gastrointestinal tract, liver, and central nervous system, which are vital for neonatal survival.2
Regulation of the HPA axis
The regulation of the fetal HPA axis and its interaction with the placenta are illustrated in fig 1. The fetal hypothalamus responds to acute stressful situations such as arterial hypotension and haemorrhage by releasing CRH. CRH stimulates the production of ACTH from the fetal corticotrophs. The latter hormone enhances adrenal cortisol secretion, which in turn inhibits excessive CRH and ACTH release from the fetal hypothalamic–pituitary centres. ACTH also promotes DHEA-S secretion from the adrenocortical fetal zone, which provides substrates for oestrogen synthesis. Placental oestrogens indirectly influence the fetal HPA axis by facilitating the conversion of active cortisol into inactive cortisone, thereby reducing the concentration of cortisol in the fetus. This results in decreasing the negative feedback effects of cortisol on the fetal hypothalamic–pituitary centres and causes an increase in fetal POMC mRNA, ACTH, and adrenal cortisol production. An increase in fetal ACTH secretion further stimulates DHEA-S production.5 This positive feedback loop has been proposed to be the mechanism underlying the rapid growth of the fetal adrenal cortex and the efficient production of DHEA-S at midgestation.5 Finally, the cortisol produced downregulates the positive feedback loop by exerting a negative control on the hypothalamic–pituitary centres (fig 1).

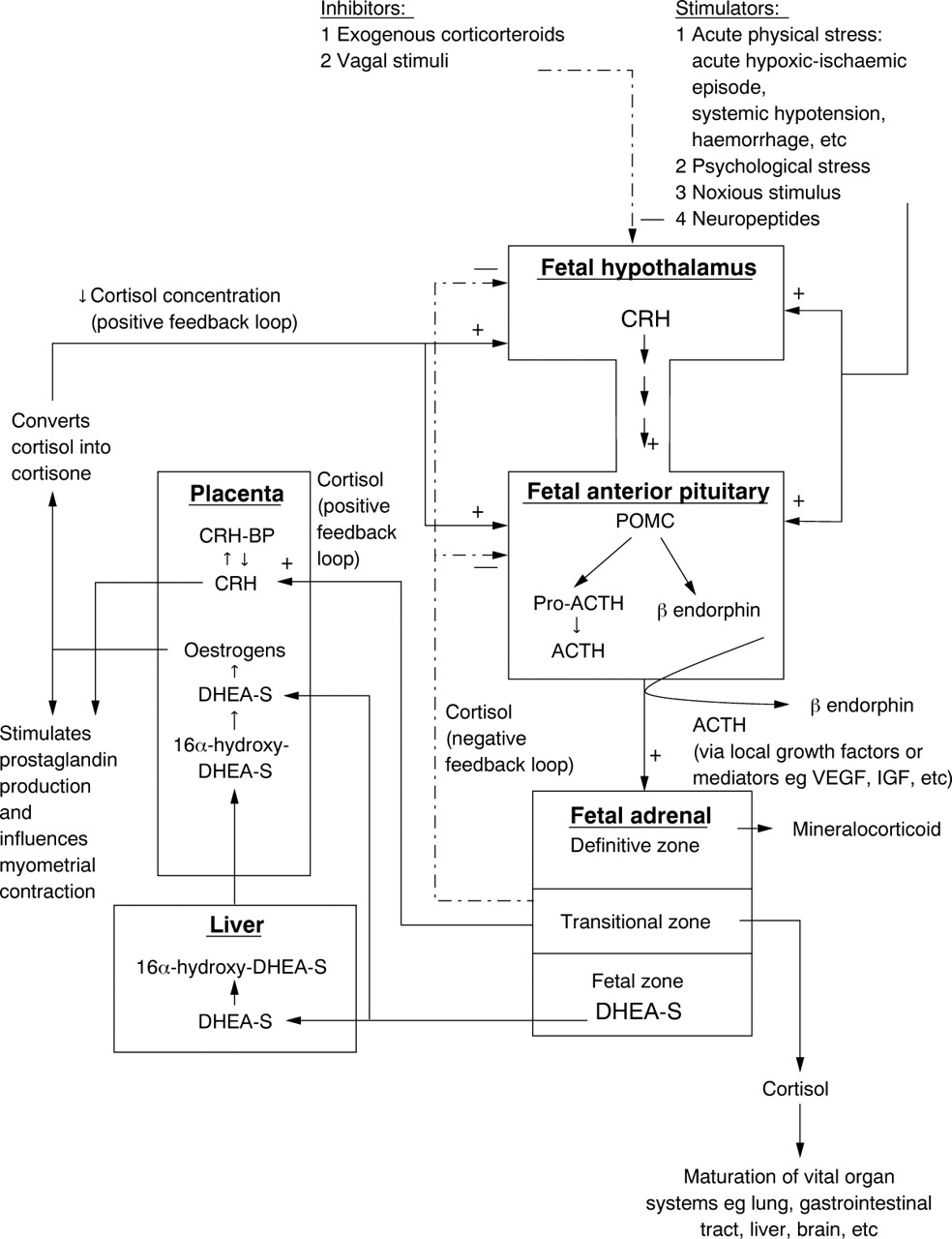{kind=link}
A schematic model to illustrate the neuroendocrine interaction between the fetal hypothalamic–pituitary–adrenal axis and the placenta. The solid arrows represent positive stimulatory pathways and the broken arrows represent negative inhibitory pathways. ACTH, corticotrophin; CRH, corticotrophin releasing hormone; CRH-BP, corticotrophin releasing hormone binding protein; DHEA-S, dehydroepiandrosterone sulphate; IGF, insulin-like growth factor; POMC, pro-opiomelanocortin; pro-ACTH, pro-corticotrophin; VEGF, vascular endothelial growth factor.
Paradoxically, adrenal glucocorticoid stimulates the release of CRH from the placenta.3 ,6 Placental CRH, which is identical to hypothalamic CRH in structure, bioactivity, and immunoreactivity, stimulates the expression of fetal pituitary ACTH and effectively establishes another positive feedback circuit, resulting in concomitant increase of CRH, ACTH, and cortisol towards the end of gestation (fig1).3 ,6 This self perpetuating mechanism reaches its peak at the onset of labour and the positive feedback loop is eventually terminated by delivery of the placenta and the fetus.3Hence, it is possible that environmental stress might stimulate a precocious rise in fetal hypothalamic and placental CRH production, which triggers the cascade of changes resulting in preterm delivery. This theory is supported by the observation that premature infants have significantly higher concentrations of maternal CRH in the early third trimester than those who deliver at term.7
Adaptation to extrauterine life
Because successful adaptation of newborns to extrauterine life relies on the exact timing of parturition and maturation of vital organ systems, the steroidogenic compartments of the fetal adrenal cortex must develop adequately and produce sufficient endogenous cortisol for perinatal survival towards the end of gestation. Despite the dramatic remodelling of the adrenal cortex that occurs immediately after birth, there is no evidence of clinical adrenocortical insufficiency in term infants during this crucial period.3In contrast, ill and extremely premature infants form a unique group because of their potentially decreased ability to produce stress induced release of glucocorticoid.8 ,9 Recent studies suggest that some preterm very low birth weight infants, in particular those less than 1000 g, have inappropriately low serum cortisol concentrations 8-10 but greatly raised concentrations of the cortisol precursors 11-deoxycortisol, 17-hydroxyprogesterone, and 17-hydroxypregnenolone when compared with term infants, indicating that the activity of some adrenal steroidogenic enzymes might be reduced as a result of adrenocortical immaturity.9 ,10 A large proportion of very low birth weight infants do not respond satisfactorily to stimulation by a physiological dose of ACTH (0.1 μg/kg).11 An inverse relation between gestational age and serum cortisol concentrations has also been described during the first 7 days of life in premature infants.12 ,13 In contrast to the above findings, Hanna and colleagues14 and Ng and colleagues15 have investigated the pituitary–adrenal function of very low birth weight infants using ovine and human CRH (1 μg/kg), respectively, and the results suggest that both ACTH and cortisol responses are adequate in most patients, with the configuration of the stimulation curves, the magnitude of responses, and the timing of the peak concentrations being comparable with those seen in the mature axis.15 Hence, it has been postulated that the relative inadequacy of cortisol production in some sick premature infants might be related to their inability to “recognise” stress or a failure of the hypothalamus to secrete CRH in stressful situations.14
Very low birth weight infants with low or suboptimal serum cortisol concentrations might have a higher risk of developing chronic lung disease and of requiring prolonged oxygen supplementation.11 ,16 Transient adrenocortical insufficiency has also been described in very low birth weight infants who present with profound hypotension and shock, which are resistant to volume expansion and inotrope treatment, but respond promptly to glucocorticoid replacement.8 In this condition, the pituitary release of ACTH in response to exogenous human CRH stimulation appears to be intact but the adrenals fail to produce adequate cortisol to maintain normal blood pressure (PC Nget al, unpublished data). This phenomenon, however, appears to be short lived and normal adrenal function returns within a month (PC Ng et al, unpublished data).8 The use of narcotic drugs for pain or stress relief should also be carefully monitored, because these agents might predispose to hypotension by lowering circulating concentrations of the stress hormones.17
The understanding of fetal and neonatal endocrine physiology has revolutionised the management of preterm, very low birth weight infants in the past decade. This knowledge has been clinically exploited by the use of antenatal and postnatal corticosteroids for advancing lung maturation and treatment of chronic lung disease.4 The effects of exogenous corticosteroids on the HPA axis are discussed in the following sections.
Antenatal corticosteroids
The use of exogenous corticosteroids for maternal or fetal indications during pregnancy has raised concerns about their potential suppressive effects on the fetal HPA axis. Transient suppression of the pituitary–adrenal glands in preterm infants whose mothers received one to two doses of antenatal corticosteroids should recover by day 7 of life.12 ,18-20 Infants exposed to corticosteroids retain their capability to respond to perinatal stress by producing appropriate amounts of cortisol.20 In addition, a recent study indicates that the pulsatile nature of adrenal secretion is also undisturbed by recent maternal corticosteroid treatment given within a seven day period before delivery.21 However, the effect of multiple courses of antenatal corticosteroid on the HPA axis is less well defined. Although our initial investigation using the human CRH stimulation test suggested that the pituitary–adrenal responsiveness of preterm infants at days 7 and 14 of postnatal life is unaffected when multiple courses (mean, 7.2 doses) of antenatal dexamethasone are administered to the mothers,12 our latest evidence indicates that mild adrenal suppression might be present in a small proportion of preterm infants whose mothers received more than eight doses (mean, 11.6).22 A significant negative correlation has also been seen between poststimulation serum cortisol concentrations and the cumulative antenatal dexamethasone doses received by the mothers.22 We can conclude from the current evidence that a standard course (two doses) of antenatal dexamethasone has no long lasting suppressive effects on the pituitary–adrenal function. Even multiple courses have relatively little clinical or biochemical influence on the HPA axis.12 ,22 The use of antenatal corticosteroids should not be withheld because of the fear of HPA axis suppression.
Postnatal corticosteroids
Recent studies have shown that the HPA axis is greatly suppressed after a three to six week course of dexamethasone.23-26Even a brief period of exposure (five to seven days) might result in severe suppression.23 ,27-29 However, a repeatable three day pulsed course of dexamethasone is associated with a lesser degree of adrenal suppression when compared with a continuous six week regimen.30 Studies using ovine CRH, human CRH, and metyrapone tests suggest that the hypothalamic–pituitary centres are also suppressed by systemic corticosteroids.24-26 ,28Despite the high dose and the prolonged course of treatment, most investigators have shown that in most infants HPA axis function recovers within one to two months after completion of the steroid course.23-26 Although our results using the human CRH stimulation test indicate that the pituitary centre is able to recover earlier than the adrenal glands,24 another study using the metyrapone test suggests that, as a functional unit, the hypothalamic–pituitary centres probably recover later than the adrenals.25 Steroid replacement treatment might be desirable at the time of stress in the immediate post-treatment period, but seems unnecessary one to two months after stopping systemic corticosteroid treatment.24 The effect of inhaled fluticasone propionate on the HPA axis has also been assessed in a randomised controlled study.31 A two week course of inhaled fluticasone propionate (1000 μg/day) produces moderately severe pituitary–adrenal suppression in very low birth weight infants.31 Inhaled corticosteroid is probably absorbed directly by the pulmonary vasculature and effectively circumvents the hepatic first pass metabolism, giving rise to systemic side effects.32 Thus, we urge vigilant surveillance in monitoring signs of HPA axis insufficiency in severely ill infants who have received postnatal systemic or high dose inhaled corticosteroids, because the pituitary and adrenal glands, although considered to be biochemically active after one to two months of stopping treatment, might have less reserve compared with their pretreatment state.23 ,24
Programming
Recent epidemiological evidence suggests that stressful events experienced in fetal and early neonatal life can produce enduring changes in the structure and function of the neural pathways, thereby resulting in alteration of the programming process which predisposes to specific diseases in later life.32-38 It has been shown that early infant separation stress is associated with an increase in adult psychopathology, and a permanent rise in β endorphin and cortisol in the circulation.39 Animal experiments on rodents further indicate that perinatal manipulation of the HPA axis by exogenous corticosteroids and stress might permanently alter the development of central monoamine neurones33 and reduce the CRH content in the median eminence, which subsequently might be associated with a decrease in stress induced CRH release in adulthood.40 The administration of exogenous corticosteroids to pregnant rats decreases the birth weight and raises the blood pressure of their offspring in later life.35 A relation between birth weight, blood pressure, glucose tolerance, and HPA axis hormones is also seen in humans. Fetuses and newborns with estimated weight and birth weight less than the 10th centile for gestational age have increased umbilical venous ACTH and cortisol concentrations when compared with larger fetuses and newborns of equivalent gestations.41 ,42 Cortisol values in adults were found to be highest in those who were lightest at birth.43 Low birth weight and raised serum cortisol concentrations are positively correlated with high blood pressure, but inversely related to glucose tolerance.35-38 ,43Therefore, it has been postulated that perinatal stress and exposure to corticosteroids might induce long lasting changes in the HPA axis and alter an individual's sensitivity to later environmental manipulations.32 ,34 Hence, the long term effects of antenatal and postnatal corticosteroids on the HPA axis and their potential to predispose to specific diseases in later life should be studied longitudinally, and carefully monitored.
Summary
Regulation of the fetal HPA axis is a highly complicated process and is under the control of positive and negative feedback circuits (fig 1), placental hormones, and local autocrine/paracrine mediators or growth factors.3 The primary aims of this complex system are to ensure appropriate coordination of tissue growth and differentiation, orderly maturation of vital organ systems, and ultimately to act together with the placenta to determine the exact timing of parturition most suitable for successful transition from intrauterine to extrauterine life. Undoubtedly, the understanding of basic fetal and neonatal neuroendocrine physiology and development has been clinically exploited and translated into new measures for revolutionising the management of preterm and newborn infants. Current evidence suggests that the antenatal and postnatal use of corticosteroids causes only transient suppression of the HPA axis, and endocrine function recovers within one week and four to eight weeks, respectively, in most infants after corticosteroid treatment has been stopped.12 ,23-26 However, because early treatment with exogenous corticosteroids might potentially influence the programming of the HPA axis,32 clinicians should monitor the long term effects of such exposure in treated cases.