Article Text

Abstract
Objective Infants with a congenital diaphragmatic hernia (CDH) are at high risk of developing pulmonary hypertension after birth, but little is known of their physiological transition at birth. We aimed to characterise the changes in cardiopulmonary physiology during the neonatal transition in an ovine model of CDH.
Methods A diaphragmatic hernia (DH) was surgically created at 80 days of gestational age (dGA) in 10 fetuses, whereas controls underwent sham surgery (n=6). At 138 dGA, lambs were delivered via caesarean section and ventilated for 2 hours. Physiological and ventilation parameters were continuously recorded, and arterial blood gas values were measured.
Results DH lambs had lower wet lung-to-body-weight ratio (0.016±0.002vs0.033±0.004), reduced dynamic lung compliance (0.4±0.1mL/cmH2O vs1.2±0.1 mL/cmH2O) and reduced arterial pH (7.11±0.05vs7.26±0.05), compared with controls. While measured pulmonary blood flow (PBF) was lower in DH lambs, after correction for lung weight, PBF was not different between groups (4.05±0.60mL/min/gvs4.29±0.57 mL/min/g). Cerebral tissue oxygen saturation was lower in DH compared with control lambs (55.7±3.5vs67.7%±3.9%).
Conclusions Immediately after birth, DH lambs have small, non-compliant lungs, respiratory acidosis and poor cerebral oxygenation that reflects the clinical phenotype of human CDH. PBF (indexed to lung weight) was similar in DH and control lambs, suggesting that the reduction in PBF associated with CDH is proportional to the degree of lung hypoplasia during the neonatal cardiopulmonary transition.
- congenital abnorm
- fetal medicine
- neonatology
- animal research
- physiology
Statistics from Altmetric.com
What is already known on this topic?
Lung hypoplasia and pulmonary hypertension associated with congenital diaphragmatic hernia (CDH) contribute to mortality rates that remain between 30% and 40% despite centralised, standardised neonatal care.
Normally at birth, lung aeration triggers an increase in pulmonary blood flow that enables gas exchange and venous return to the left ventricle via the pulmonary circulation.
Infants with CDH have small pulmonary vascular cross-sectional area and increased distal vessel muscularisation. These structural changes likely influence cardiopulmonary physiology during the neonatal transition.
What this study adds?
Lung aeration stimulated a large increase in pulmonary blood flow (PBF) that was proportional to lung size in both DH and control lambs.
PBF indexed to lung weight was similar between DH and control lambs, indicating that the lower PBF and higher pulmonary vascular resistance seen in DH lambs are mostly because the lung is smaller.
During hypoxaemia, titrating fraction of inspired oxygen based on oxygen saturation (SpO2) causes an overshoot in cerebral oxygen delivery due to increased carotid blood flow.
Introduction
Congenital diaphragmatic hernia (CDH) is a rare birth defect (1 in 2500 live births) caused by the failed closure of the diaphragm in the embryonic period.1 CDH allows visceral organs to herniate into the fetal chest, which disrupts lung development. Though surgically correctable, neonates with CDH face significant respiratory challenges after birth.2 The lungs are smaller and structurally abnormal in both airway and vascular development, resulting in respiratory insufficiency immediately after birth that is often associated with pulmonary hypertension.3–8 Despite early referral to high volume care centres for intensive neonatal care, neonatal mortality of isolated CDH remains high (30%).2 9–11
Standardised neonatal care for CDH infants is largely based on expert opinion, and outside of clinical trials, antenatal management is expectant.8 12 To further improve antenatal and neonatal management of CDH infants, we require a better understanding of how CDH affects the physiological transition at birth.
The transition to newborn life at birth involves the lungs taking over the role of gas exchange from the placenta and the closure of cardiovascular shunts to separate the pulmonary and systemic circulations. Critical to these cardiopulmonary changes is a reduction in pulmonary vascular resistance (PVR), which enables the lung to accept the entire output of the right ventricle while also facilitating a reduction in pulmonary arterial pressure (PAP). However, development of the pulmonary vasculature is impaired in infants with CDH, resulting in a small cross-sectional area predominantly in the ipsilateral lung.4 Combined with this, infants with CDH have abnormal thickening of the smooth muscle surrounding small pulmonary arteries. This abnormal development, together with an increased sensitivity to vasoactive triggers such as hypoxia, are thought to contribute to the maintenance of a high PVR in the newborn period, leading to the development of persistent pulmonary hypertension of the newborn (PPHN). However, it remains unclear what drives this abnormal vascular development and whether maladaptation to the cardiopulmonary changes associated with birth may contribute to the development of PPHN.
Birth-related cardiopulmonary adaptations are not well described in infants with CDH, particularly the capacity for PVR to decrease in response to lung aeration. We aimed to describe the cardiopulmonary physiology during transition to neonatal life in lambs with pulmonary hypoplasia due to a diaphragmatic hernia (DH). In DH rabbit kittens, we have recently found that the increase in pulmonary arterial blood flow (PBF) induced by lung aeration in the normal lung also occurs in the hypoplastic lung.13 Therefore, in this study, we hypothesised that abnormal lung development may predispose DH lambs to low lung compliance and respiratory insufficiency, but any reduction in pulmonary blood flow during the transition period will be proportional to the reduction in lung size.
Methods
Animal surgery and treatment groups
All surgical procedures were performed under general anaesthesia using intravenous sodium thiopental (20 mg/kg, Pentothal; Jurox, New Zealand) for induction and inhaled isoflurane (≈2% in room air; Isoflow, Abbot) for maintenance. Ewes were intubated and monitored (ECG, expired CO2 and oxygenation) during surgery.
After surgery, the ewes received 3 days of analgesia (transdermal fentanyl patch, 75 µg/hour; Janssen Cilag) and were monitored daily until delivery. Before surgery, animals were allocated to control or DH groups.
Ultrasound evaluation
Before every procedure, a fetal ultrasound was performed (CX50 Ultrasound System, Philips) to measure the thoracic and cardiac circumference, right and left lung area, midtracheal diameter and femur length. The shape of the fetal skull and limited visualisation at later gestational ages precluded reproducible images of the fetal head. We calculated the right lung to femur ratio (RLFR).
DH creation
A DH was surgically created at ≈80 days of gestational age (dGA; term≈147 dGA). The ewe was given cefazolin (intravenous) (1 g, AFT Pharmaceuticals) before a midline laparotomy was used to expose the uterus. The fetal head and forelimbs were exteriorised via hysterotomy, taking care to avoid placental cotyledons. The fetal diaphragm was visualised via a thoracotomy through the ninth intercostal space, and the diaphragm was incised and resected, enabling the stomach and bowels to be repositioned into the fetal chest cavity. The thoracotomy incision was sutured closed (Maxon 2–0; Covidien) before the fetus was returned to the amniotic sac, and the uterus was closed in two layers (polysorb 2–0, Covidien); leaked amniotic fluid was replaced using warmed saline, and intra-amniotic cefazolin (1 g) was given. The maternal fascia, subcutaneous tissue and the skin were then closed (Maxon 0, Covidien and Vetafil 2–0; Supramid). In control fetuses the thorax was opened, but there was no incision of the diaphragm.
Delivery and ventilation
At ≈138 dGA, the ewe was anaesthetised (as above), and the fetal head and neck were exposed via hysterotomy. The lamb was intubated using a size 4.0 cuffed endotracheal tube. Polyvinyl catheters were inserted into the jugular vein and carotid artery to allow neonatal drug administration and continuous carotid arterial pressure monitoring, respectively. An ultrasonic flow probe (Transonic Systems, Ithaca, New York, USA) was placed around the contralateral carotid artery to continuously record carotid arterial blood flow (CBF). The fetal chest was exteriorised, and a thoracotomy was performed through the fourth intercostal space. An ultrasonic flow probe was placed around the left pulmonary artery to continuously record PBF. A non-occlusive catheter was implanted in the main pulmonary artery, which was connected to a pressure transducer to continuously record PAP. The thorax was closed (Maxon 2–0) to prevent air leakage during ventilation. Pulse-oximeter, near-infrared spectroscopy (NIRS) and temperature probe were placed on the right forelimb, skull and rectally, respectively. The stomach was drained via an orogastric tube. After instrumentation, the lamb was delivered from the uterus, dried and carefully positioned on the ewe’s abdomen to avoid obstruction of umbilical cord blood flow.
After the lung liquid was passively drained, ventilation commenced with a 30 s sustained inflation (35 cmH2O, 21% O2) followed by intermittent positive pressure ventilation (iPPV) in volume guarantee mode using tidal volume of 4 mL/kg (Babylog 8000, Dräger, Lübeck, Germany). Positive end-expiratory pressure was set at 5 cmH2O and peak inspiratory pressure (PIP) was initially limited to 35 cmH2O. If the target tidal volume was not reached after 30 s, a second sustained inflation was performed followed by iPPV for a total of 120 min. The umbilical cord was clamped when a tidal volume of 4 mL/kg was achieved or at 10 min after ventilation onset. The lamb was then moved to a warming bed, sedated with continuous alfaxalone (10 mg/mL, 5 mL/hour; Alfaxan, Jurox) and covered with plastic drapes to maintain a stable body temperature. Lambs were ventilated with warmed and humidified oxygen. Ventilator rate, inspiratory and expiratory rate and fraction of inspired oxygen (FiO2) were titrated to achieve gas targets for arterial carbon dioxide tension (PaCO2) 60–80 mm Hg, arterial oxygen tension (PaO2) >40 mm Hg and oxygen saturation (SpO2) of 85%–88%. The lamb was euthanised (sodium pentobarbitone intravenous 100 mg/kg) at the end of the 2-hour ventilation protocol or earlier in case of significant pneumothorax or severe acidosis unresponsive to alteration of ventilation parameters.
Outcome measures
Arterial blood samples were analysed every 5 min during the first 30 min of ventilation and every 10 min thereafter.
Physiological data analysis
Tidal volume, airway pressures, cerebral tissue oxygen saturation (SctO2), pulmonary and cerebral perfusion were continuously recorded using LabChart (ADInstruments, New South Wales, Australia) and analysed offline. Ten heartbeat average data points were collected prior to ventilation onset (fetal), immediately at the onset of ventilation (t=0) and every 5 min throughout the ventilation period.
Postmortem examination
The presence of a diaphragmatic defect and herniation of visceral organs was confirmed during postmortem examination. The lungs were weighed and expressed as a ratio to the body weight (wet lung-to-body weight ratio [LBWR]). Lung size (LBWR) was used as a proxy for degree of lung hypoplasia, as it has previously been shown to also correlate with radial alveolar count and lung DNA content in a similar ovine CDH model.14
Calculations
PVR (mm Hg/[mL/min]): (pulmonary artery pressure – left atrial pressure)/pulmonary blood flow. Based on previous studies, the left atrial pressure was assumed to equal 9 mm Hg.15
Alveolar-arterial difference in oxygen tension (AaDO2; mm Hg): (FiO2*713−PaCO2/0.8)−PaO2.
Cerebral oxygen extraction (%): (SaO2−SctO2)/SaO2, where SaO2 is the arterial oxygen saturation.16
Arterial oxygen content (CaO2; mL O2/L): [1.39·Hb·SaO2/100] + [0.03·PaO2]), where Hb is the haemoglobin concentration (g/L).17
Cerebral oxygen delivery (DO2; mL/min/kg): (CBF·CaO2).17
Statistical analysis
Based on power analysis from previous studies, we estimated ≈7 animals per group and were required to detect minimal biologically significant differences in PBF and ventilation characteristics with a coefficient of variation of ≈25%, assuming an α value of 0.05 and a power of >0.8.18 Continuous data are expressed as means±SEM or medians (IQR) if the data were not normally distributed. Dichotomous data are expressed as percentages. Differences between control and DH groups were analysed using two-way repeated measures analysis of variance combined with Holm-Sidak’s multiple comparisons test. A p value below 0.05 was considered statistically significant.
Results
Animal surgery
Thirteen (of 16) fetuses survived to delivery: six controls (100%) and seven DH lambs (70%). All control lambs survived the 2-hour ventilation period; however, for two DH lambs, it was necessary to discontinue the experimental protocol early due to developing treatment-resistant pneumothoraces (at 70 min and 110 min).
Postmortem examination
A diaphragmatic defect with substantial gastrointestinal herniation into the thorax was confirmed at postmortem examination in all DH lambs. The wet LBWR for DH animals was significantly smaller than controls (0.016±0.002 vs 0.033±0.004; p=0.02).
Ultrasound
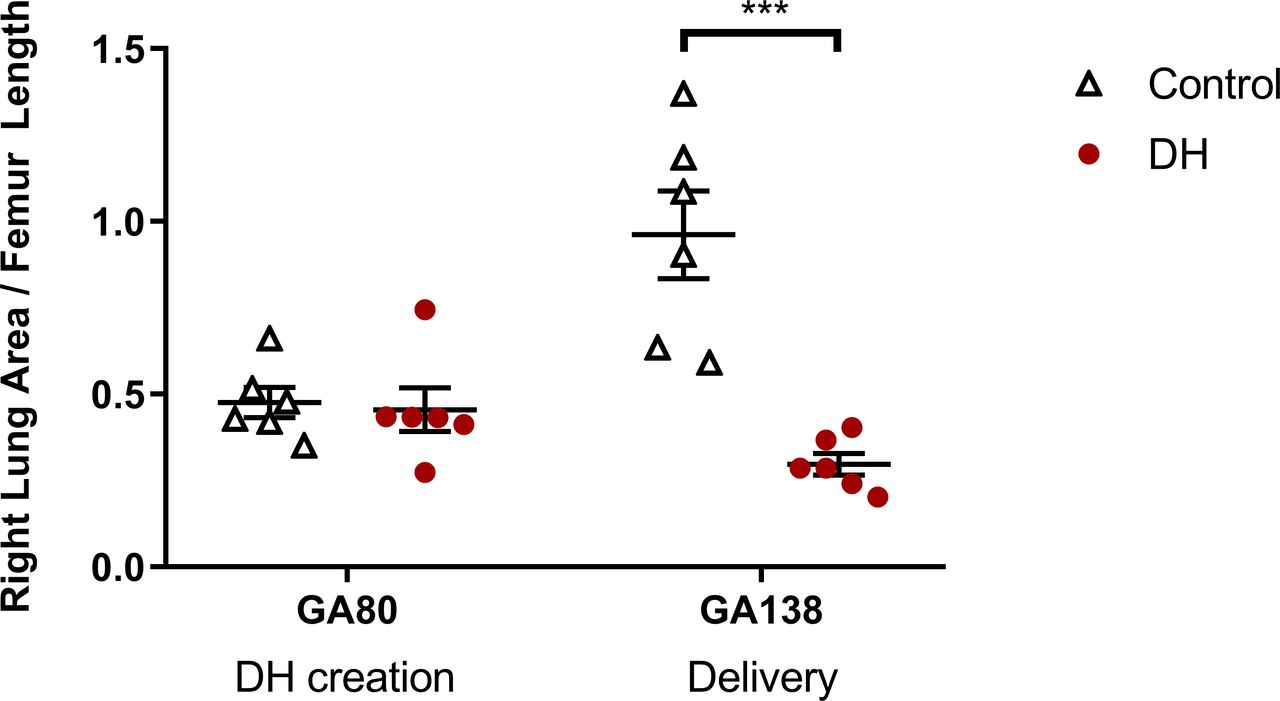
In control fetuses, RLFR increased between sham surgery at ≈80 dGA and delivery at ≈138 dGA (0.48±0.04 vs 0.96±0.13; p=0.002, figure 1). In contrast, RLFR in DH fetuses decreased between DH creation at ≈80 dGA and delivery at ≈138 dGA (0.46±0.06 vs 0.30±0.03; p=0.044).
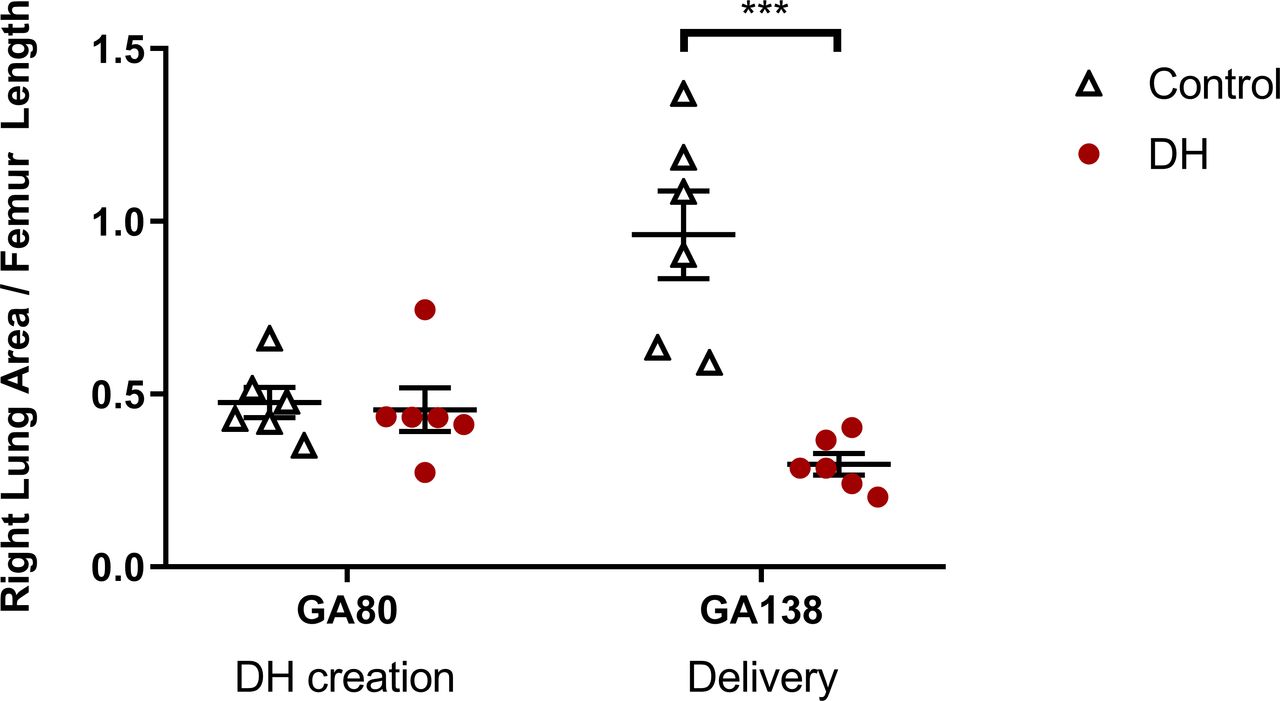
The ratio of fetal right lung area to femur length assessed using ultrasound throughout gestation. Groups shown are control (white triangles, n=6; thoracotomy only at ≈80 dGA, delivery at ≈138dGA) and diaphragmatic hernia (DH) (red circles, n=7; DH creation at ≈80 dGA, delivery at ≈138 dGA). Presented as mean±SEM and significance accepted when p<0.05. Asterisks (***) indicate significant difference (p<0.001) between control and DH at delivery. dGA, days of gestational age.
Ventilation and oxygenation
Only two DH lambs reached the target tidal volume (VT; 4 mL/kg) within the 120 min experiment (at 20 min and 25 min), whereas control lambs reached target VT at approximately 5 min (IQR 5–6.25). Hence, the umbilical cord was clamped at t=10 min in DH lambs and t≈5 min in control lambs. To obtain these tidal volumes, DH lambs required a higher PIP than controls (36.9±2.3 cmH2O vs 25.3±2.5 cmH2O; p=0.009); therefore, dynamic lung compliance was significantly lower in DH lambs throughout the 2-hour ventilation period (0.4±0.1 mL/cmH2O vs 1.2±0.1 mL/cmH2O; p=0.001, figure 2A).

(A) Dynamic lung compliance, (B) partial pressure of arterial carbon dioxide (PaCO2), (C) alveolar-arterial difference in oxygen tension (AaDO2) and (D) arterial oxygen saturation (SaO2) over the 2 hours following delivery at ≈138 dGA. Groups shown are control (white triangles, n=6; thoracotomy only at ≈80 dGA) and diaphragmatic hernia (DH) (red circles, n=7; DH creation at ≈80 dGA). Presented as mean±SEM and significance accepted when p<0.05. Asterisk (*) indicates significant differences between groups (*p<0.05, ***p<0.001) at all individual timepoints below line. dGA, days of gestational age.
Before ventilation onset, DH and control lambs had similar PaCO2 and arterial pH and these parameters remained stable in both groups during the initial period of ventilation with an intact umbilical cord (figure 2A). However, in DH lambs, PaCO2 increased and arterial pH decreased immediately after the umbilical cord was clamped (t=10 min). In DH lambs, by 40 min after ventilation onset, PaCO2 was significantly higher (102.0±16.0 vs 57.2±3.4 mm Hg; p=0.029) and pH lower (7.08±0.07 vs 7.28±0.02; p=0.017) than controls, and these differences persisted for the remainder of the 2-hour ventilation period (figure 2B).
The AaDO2 was similar between DH and control lambs, while the umbilical cord remained intact (figure 2C). However, the AaDO2 became markedly increased in DH lambs compared with controls immediately after the umbilical cord was clamped (485±74 mm Hg vs 129±37 mm Hg; p<0.001). Despite having a high FiO2 throughout the 2-hour ventilation period (0.86±0.08 vs controls 0.36±0.08; p<0.001), DH lambs had a lower SaO2 than controls from 80 min after ventilation onset and onwards (66.1±12.3 vs 90.7%±1.0%; p=0.025, figure 2D).
Pulmonary perfusion
Mean PBF increased rapidly in both groups, reaching a maximum at 20 min following ventilation onset. The rate of PBF increase (2.06 mL/min/kg per min vs 5.57 mL/min/kg per min; p<0.001) and the maximum PBF reached (41.8±9.1 mL/min/kg vs 94.6±10.8 mL/min/kg; p<0.001) were lower in DH lambs compared with controls. Following this maximum, PBF levels stabilised and remained relatively constant although tended to gradually decrease in both groups during the remainder of the 2-hour ventilation period (figure 3A). At 2 hours, PBF was significantly lower in DH compared with control lambs (21.4±7.2 mL/min/kg vs 50.5±12.7 mL/min/kg; p=0.048).

Pulmonary blood flow indexed to (A) body weight and (B) left lung weight, and pulmonary vascular resistance indexed to (C) body weight and (D) left lung weight over the 2 hours following delivery at ≈138 dGA. Groups shown are control (white triangles, n=6; thoracotomy only at ≈80 dGA) and diaphragmatic hernia (DH) (red circles, n=7; DH creation at ≈80 dGA). Presented as mean±SEM and significance accepted when p<0.05. Asterisk (*) indicates significant differences between groups (**p<0.01, ***p<0.001) at all individual timepoints below line. dGA, days of gestational age.
After correcting for lung weight, the increase in PBF was similar in DH and control lambs over the 2-hour ventilation period, although the rate of increase was slower in DH lambs. PBF (per gram of lung weight) was lower at 10 min (2.95±1.06 mL/min/g vs 5.83±0.54 mL/min/g; p=0.009) after ventilation onset in DH lambs compared with controls but not significantly different at any other timepoint (figure 3B).
PAP was similar to systemic arterial pressure in both groups before ventilation onset (figure 4A). In control lambs, by 10 min after ventilation onset PAP was 3.3±1.8 mm Hg lower than systemic arterial pressure and by 60 min was 11.7±3.2 mm Hg lower. In contrast, PAP remained similar to systemic arterial pressure throughout the 2-hour experiment in DH lambs (figure 4A).
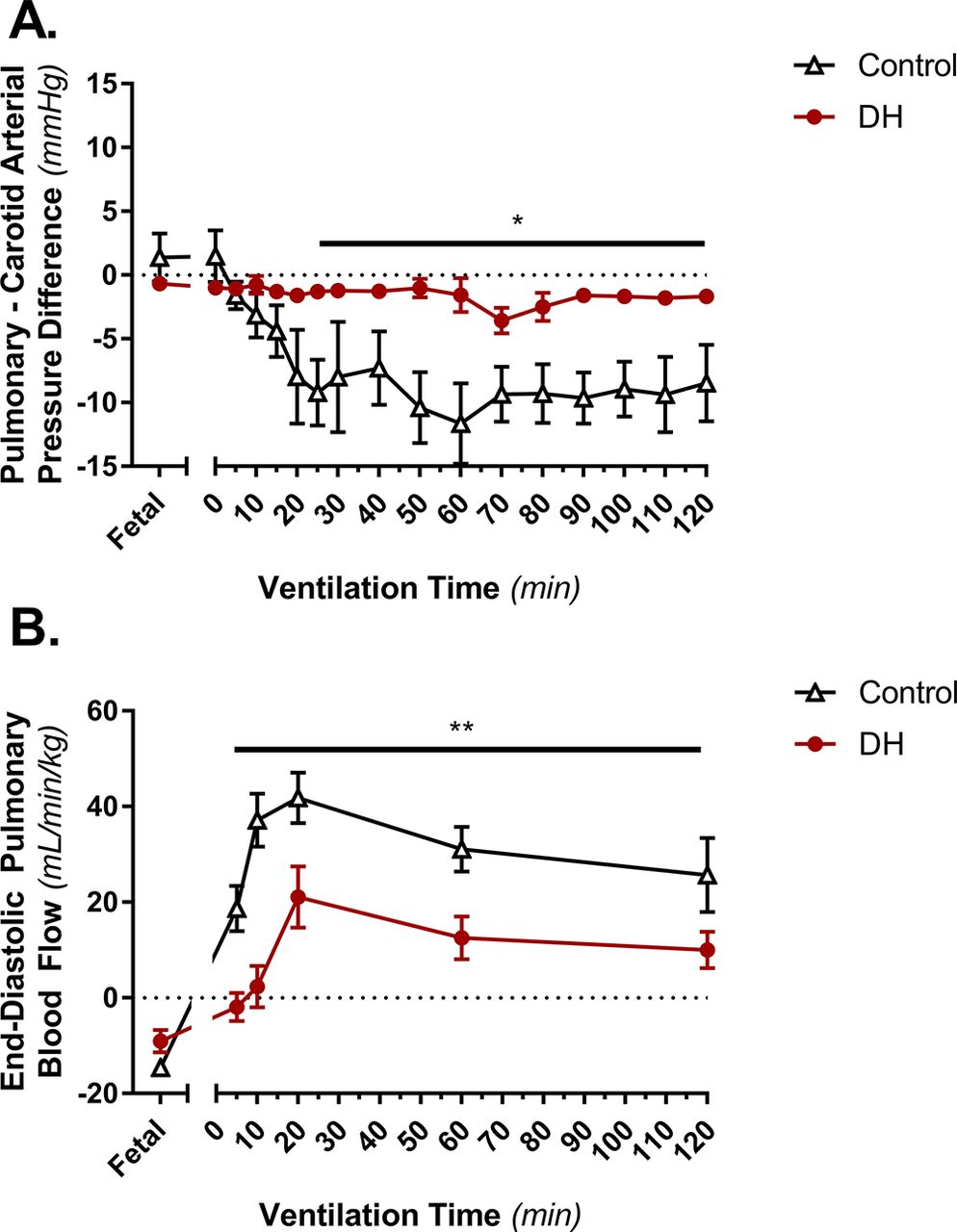
(A) The difference between pulmonary arterial pressure and systemic arterial pressure and (B) end-diastolic pulmonary blood flow over the 2 hours following delivery at ≈138 dGA. Groups shown are control (white triangles, n=6; thoracotomy only at ≈80 dGA) and diaphragmatic hernia (DH) (red circles, n=7; DH creation at ≈80 dGA). Presented as mean±SEM and significance accepted when P<0.05. Asterisk (*) indicates significant differences between groups (*p<0.05, **p<0.01) at all individual timepoints below line. dGA, days of gestational age.
End-diastolic PBF was retrograde (assigned a negative value) in both groups before ventilation onset (figure 4B), which is a normal characteristic of PBF in the fetus.19 At 5 min after ventilation onset, when the umbilical cord had been clamped in controls but not in DH lambs, end-diastolic flow was positive in controls but remained mostly retrograde in DH lambs (18.7±4.8 mL/min/kg vs −1.9±3.0 mL/min/kg; p=0.004). After umbilical cord clamping, end-diastolic flow was positive in both groups throughout the 2-hour experiment; however, it was greater in controls compared with DH lambs (at 2 hours 25.7±7.7 mL/min/kg vs 10.0±3.8 mL/min/kg; p=0.024).
PVR was high in both DH lambs and controls before ventilation onset (3.37±1.30 vs 2.07±0.77 mm Hg/(mL/min); p=0.068) but markedly decreased after ventilation onset in control lambs. In contrast, in DH lambs PVR did not decrease until 15 min after ventilation onset. As such, at 5 min after ventilation onset PVR was significantly greater in DH lambs compared with controls (1.95±0.55 vs 0.30±0.05 mm Hg/(mL/min); p=0.014). While PVR did eventually decrease in DH lambs (figure 3C), from 15 min to 120 min, it remained greater than in controls (0.53±0.06 vs 0.20±0.05 mm Hg/(mL/min); p=0.003). After correcting for lung weight, PVR was not different between DH and control lambs at any timepoint (figure 3D).
Cerebral perfusion
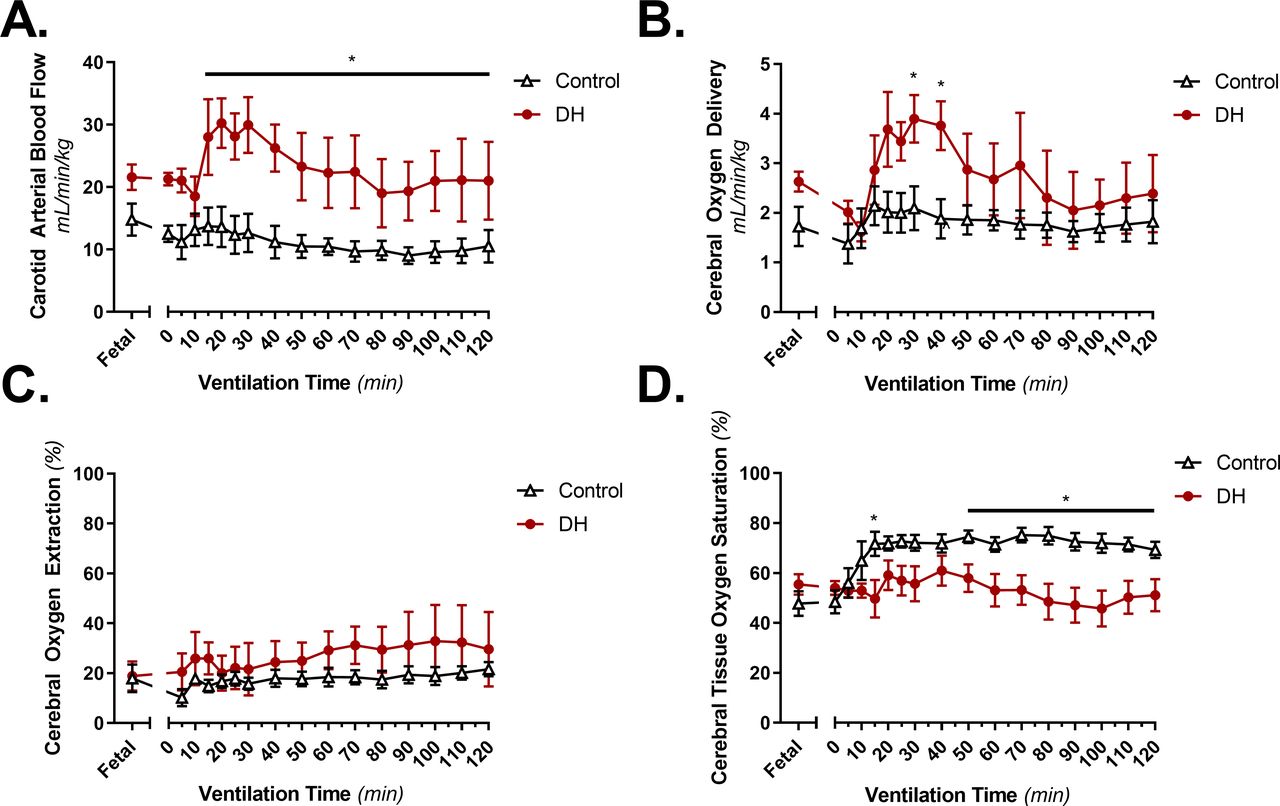
Cerebral DO2 was similar in DH lambs and controls before ventilation onset. In control lambs, DO2 remained at ≈2 mL/min/kg throughout the 2-hour experiment (figure 5B). Despite the persistently low SaO2 described above, in DH lambs, DO2 initially increased to a maximum at 30 min (3.9±0.5 vs controls 2.1±0.5 mL/min/kg; p=0.015) before returning to similar levels as controls by the end of the 2-hour experiment (figure 5B). Underlying this, in DH lambs CBF initially increased to a maximum at 30 min (30.0±4.5 mL/min/kg vs controls 12.7±3.1 mL/min/kg; p=0.002) and remained greater than controls throughout the 2-hour experiment (at 2 hours 21.0±6.2 mL/min/kg vs 10.5±2.6 mL/min/kg; p=0.038, figure 5A). Cerebral oxygen extraction was not significantly different between groups throughout the 2-hour neonatal ventilation (figure 5C). SctO2 was not significantly different between groups during the first 60 min of ventilation; however, towards the end of the experiment, it was lower in DH lambs compared with controls and remained lower until the end of the experiment (figure 5D).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Carotid blood flow (mL/min/kg), (B) cerebral oxygen delivery (mL/min/kg), (C) cerebral oxygen extraction (%) and (D) cerebral tissue oxygen saturation (%) over the 2 hours following delivery at ≈138 dGA. Groups shown are control (white triangles, n=6; thoracotomy only at ≈80 dGA) and diaphragmatic hernia (DH) (red circles, n=7; DH creation at ≈80 dGA). Presented as mean±SEM and significance accepted when p<0.05. Asterisk (*) indicates significant differences between groups (p<0.05) at all individual timepoints below line. dGA, days of gestational age.
Discussion
Using this ovine CDH model, we can invasively monitor and directly measure the cardiopulmonary changes that are responsible for the clinical challenges associated with supporting an infant with CDH through the neonatal transition. Our DH lambs demonstrated reduced dynamic lung compliance, low tidal volumes and respiratory acidosis that closely reflect the clinical experience of supporting CDH infants after birth.20 Following lung aeration, DH was associated with increased PAP compared with controls, and despite higher input pressures, PBF was decreased. Low PBF and high PAP is indicative of a high PVR that likely contributes to impaired gas exchange and subsequent hypoxaemia and hypercapnia observed in DH lambs and also in infants with CDH.2 However, after correcting for lung weight, we found that PBF was similar in control and DH lambs. These observations are similar to those reported by Hill et al,21 who observed a decrease in PBF proportional to the degree of lung hypoplasia in fetal lambs with DH. Thus, the primary mechanism for the higher PVR and lower uncorrected PBF in DH lambs seems mostly due to a smaller lung rather than an abnormal vasodilatory response. This result is also consistent with our previous finding in DH rabbits and indicates that any pulmonary vessel smooth muscle hypertrophy and neomuscularisation that may have occurred prenatally does not directly impact on the capacity of the vessels to vasodilate in response to lung aeration.13
It is interesting that O’Toole et al 22 previously observed that, when corrected for lung weight, DH lambs had reduced PBF levels compared with controls following ventilation onset. Compared with our DH lambs, their DH lambs were generally more hypoxic, hypercapnic and acidotic despite the DH being created at a similar timepoint (≈78 dGA). One methodological difference between the two studies relates to the timing of umbilical cord clamping. We clamped the umbilical cord after achieving our target tidal volume, whereas they clamped the umbilical cord immediately before ventilation commenced. This methodological difference could explain the discrepancy between the two experiments because immediate cord clamping is known to cause a rapid (within four heart beats) and large (30%) increase in afterload for the heart. Due to the presence of an open ductus arteriosus (DA), this increased afterload is experienced by both the left and right ventricles and must also influence PA pressure. However, cord clamping after the lung has aerated and PBF has increased greatly reduces this increase in afterload, because the pulmonary circulation becomes an alternative low resistance pathway for blood to flow via left to right shunting through the DA.23 24 While this characteristic is reliant on a reduction in PVR, significant left-to-right shunting through the DA was observed in all DH lambs following lung aeration, as indicated by forward flow into the pulmonary artery throughout diastole (figure 4B). Thus, although the reduction in PVR associated with lung aeration was delayed and diminished in DH lambs, by also delaying cord clamping there was sufficient time for PVR to fall, allowing left-to-right DA shunting and thereby minimising the increase in ventricular afterload caused by cord clamping. These findings suggest that the effect of delayed umbilical cord clamping in infants with CDH certainly warrants further research in experimental models, and feasibility studies for clinical trials have already commenced.25
While PBF increased in DH lambs after ventilation onset, the increase occurred later than controls, and the rate of increase was much slower. This finding could be the result of impaired vasoreactivity; however, we speculate that the greatest contribution is resulting from the slower rate of lung aeration due to poor lung compliance. We anticipated this slower rate of lung aeration and tried to mitigate the difference by initiating ventilation with a sustained inflation, as it has been shown to aerate the lungs more rapidly and uniformly and to increase PBF during the transition at birth.26
Normally after birth, CBF gradually decreases in response to an increase in oxygenation, which is an autoregulatory process that has been well described.27 28 In contrast, we observed a marked increase in CBF in our DH lambs immediately after cord clamping as they rapidly became hypoxaemic after losing placental circulatory support. This increase in CBF helped to maintain adequate DO2; however, at the same time, we were increasing FiO2 in response to rapidly deteriorating SpO2. Together, the autoregulatory increased cerebral blood flow and iatrogenically improved oxygenation led to a dramatic overshoot in oxygen delivery to the brain: DO2 was twofold greater in DH lambs for 30 min of the 2-hour experiment, which may have predisposed these lambs to hyperoxia-induced brain injury. Clearly, clinicians must consider autoregulatory responses to hypoxaemia during the neonatal transition before increasing FiO2 in response to low SpO2, and management may be better guided by the use of NIRS to directly assess cerebral oxygenation.
Our ovine CDH model demonstrates features of severe lung hypoplasia and pulmonary hypertension that are consistent with human CDH, and we report the physiology underpinning these clinical features during the fetal to neonatal cardiopulmonary transition. By understanding these physiological changes, we can better target antenatal and neonatal interventions. A potential limitation of our study is that the DH was surgically induced during the late pseudoglandular/early canalicular stage of lung development. In humans, the potential for pulmonary development to be disrupted starts from the pseudoglandular stage following failure of the diaphragm to close during the embryonic stage. As such, the effects of CDH on vascular and airway development may commence at an early stage when the larger airways and vessels are forming. Nevertheless, we observed significantly reduced LBWR, reduced dynamic lung compliance and impaired cardiopulmonary physiology in DH lambs that is consistent with that observed in humans. While our study is limited by the lack of histological data, Pringle et al 29 described both hypoplastic lung morphology and less prominent alveolar capillaries even with a DH creation at ~78 dGA. However, as diaphragmatic defects were created at 80 dGA, the effects on lung airway and vascular development may be less profound than when the DH is induced at ~63 dGA.30
In summary, we have unearthed novel findings related to the changes in PBF during the first 2 hours after birth that reshape our understanding of how infants with CDH may transition to life after birth. Despite small, non-compliant lungs and respiratory acidosis, DH lambs experienced an increase in PBF following lung aeration of a similar magnitude to control lambs after correcting for lung weight. We now aim to use this model to investigate antenatal surgical and medical therapies that aim to rescue the lung from hypoplasia, and directly measure their effects on the neonatal cardiopulmonary transition.31
References
Footnotes
Contributors All authors included on this paper fulfil the criteria of authorship, specifically. PLJD, KJC, JAD, SBH, MT and RJH designed the experiments. PLJD, KJC, AJK, SMS, MT, SBH, JAD and RJH were essential for establishing the model. AJK, PLJD, KJC and SH were responsible for data analysis. AJK, KJC and PLJD wrote the first draft of the manuscript. All authors contributed by modifying and editing the manuscript and all approved final version.
Funding This research project was funded by grants from the Royal Australian and New Zealand College of Obstetricians and Gynaecologists Foundation, Cabrini Foundation, CDH Australia, and the Victorian Government’s Operational Infrastructure Support Program.
Disclaimer These funders were not involved in the study design; in the collection, analysis and interpretation of the data; in the writing of the report; or in the decision to submit the paper for publication.
Competing interests None declared.
Ethics approval The experiment was performed in accordance with guidelines established by the National Health and Medical Research Council of Australia. This study was approved by the relevant animal ethics committee at Monash University.
Provenance and peer review Not commissioned; externally peer reviewed.
Patient consent for publication Not required.