Article Text

Abstract
Objective Preterm infants have relative adrenal and kidney immaturity. Recently, we linked their urine sodium loss to a hypoaldosteronism at variance with an appropriate stimulation of the renin-angiotensin system. To investigate this defective aldosterone secretion, we analyse the biosynthesis pathways of adrenal steroids in neonates according to gestational age (GA).
Design Multicentre study (Premaldo) including 152 neonates classified into three groups: group 1 (very preterm (VPT)): <33 gestational weeks (GW); group 2 (preterm (PT)): 33–36 GW and group 3 (term (T)): ≥GW.
Method Steroidomic profiles of mineralocorticoids, glucocorticoids and adrenal androgens were established from umbilical cord at birth (n=152) and peripheral blood at day 3 (n=70) using a recently developed liquid chromatography mass spectrometry method (LC-MS/MS). The enzymatic activity of each biosynthesis step was estimated by the product-to-substrate ratio.
Results At birth, VPT infants exhibit a global defect in adrenal steroid synthesis pathways leading to lower levels of aldosterone, cortisol and androstenedione than in term infants. This defect was strongly related to GA. On day 3, steroid precursors (progesterone, 11-deoxycorticosterone (DOC), 17-hydroxyprogesterone(17-OH-P) and 11-deoxycortisol (S)) were higher in VPT and negatively correlated with GA. Despite of precursors’ accumulation, aldosterone and cortisol were similar in the three groups. At birth and day 3, a low cortisol/11-deoxycortisol ratio was found in preterm infants, suggesting an 11-beta-hydroxylase activity (CYP11B1) deficiency.
Conclusions At birth, VPT infants exhibit a global deficit in mineralocorticoids, glucocorticoids and adrenal androgens that attenuates on day 3 of life. Steroid profiling using LC-MS/MS provides evidence for a partial defect in 11-hydroxylase along with prematurity.
- prematurity
- steroid profiling
- mineralocorticoids
- glucocorticoids
- adrenal cortex
Statistics from Altmetric.com
What is already known on this topic?
The high urine sodium loss at birth in preterm infants is linked to a hypoaldosteronism at variance with an overactivation of the renin–angiotensin system.
Very little is known about the adrenal steroid biosynthesis pathways in preterm infants during the first days of life.
To precise the underlying mechanisms of this deficit, LC-MS/MS analyses of mineralocorticoids, glucocorticoids and adrenal androgens were performed in neonates according to gestational age.
What this study adds?
At birth, a global defect of adrenal steroid pathways leads to reduced levels of aldosterone, cortisol and androstenedione in preterm infants.
On day 3, circulating steroid precursors accumulate in blood of very preterm infants while aldosterone and cortisol levels return to normal values.
Steroid profiling provides additional support for a partial defect in 11-hydroxylase activity in very preterm infants.
Introduction
Preterm (PT) infants have an inability to maintain hydroelectrolytic homeostasis related to inappropriate renal responses to corticosteroid hormones.1 2 The mature adrenal cortex is divided in three distinct zones that produce a complex array of adrenocortical steroid hormones: mineralocorticoids in the zona glomerulosa (ZG), glucocorticoids in the zona fasciculata (ZF) and adrenal androgens in the zona reticularis (ZR).3 Recently, Gomez-Sanchez et al demonstrated a zone-specific location of the steroidogenic enzymes CYP11B1 in ZF and CYP11B2 in ZG, involved in the final steps of biosynthesis of cortisol and aldosterone, respectively, while CYP11A, HSD3β and CYP21A2 are all expressed in both zonae.4 In fetal adrenal cortex, these three zones become morphologically distinguishable during the third trimester of pregnancy. The inner or fetal zone expresses CYP11A1, CYP17 and sulfokinase activities and produces dehydroepiandrosterone (DHEA) and dehydroepiandrosterone sulfate (DHEAS).5 In the transitional zone, CYP11A1, CYP17, CYP21A2 and CYP11B1 support cortisol production. Enzymes required for aldosterone synthesis, such as CYP21A2 and CYP11B2, are not expressed in the outer definitive zone during early stages of gestation.6 Thus, this zone acquires the capacity to synthetize mineralocorticoids during late gestation. After birth, the fetal zone involutes and disappears at approximately 6 months of age and the definitive and transitional zones fully differentiate into ZG and ZF by the age of 3 years.3
These temporal and spatial variations in steroidogenic enzyme expression during late gestation may affect the steroid secretion in PT infants. The human fetal adrenal cortex is unable to produce cortisol de novo until 30 gestational weeks (GW).7 Several studies have reported a relative hypocortisolism after preterm birth and suggested an immaturity of adrenal 21- and/or 11β-hydroxylase activities in premature infants.8–11 This relative adrenocortical insufficiency was associated to a low outcome and an increased morbidity such as bronchopulmonary dysplasia.12 13
In contrast, very few studies have investigated the mineralocorticoid pathway during the neonatal period. We previously reported that term neonates exhibit at birth a partial aldosterone resistance associated with undetectable renal mineralocorticoid receptor expression. This coincides with transient urinary sodium loss and altered ability to reabsorb sodium and water.14–16 Recently, the Premaldo study highlighted a deficiency in aldosterone secretion at birth in very preterm (VPT) infants. This defect was accompanied by a stimulation of the renin–angiotensin system, suggesting a partial alteration of the mineralocorticoid pathway.17
To investigate the underlying mechanisms and evaluate the impact of prematurity on global steroid hormone production, mineralocorticoid and glucocorticoid profiles were assayed during the first days of life in PT and term (T) infants. For that purpose, we used a highly sensitive and specific LC-MS/MS method that enables a full exploration of adrenal pathways including minor steroids and allows an indirect evaluation of the biosynthesis enzyme activities.
Material and methods
Patients
Umbilical cord blood samples of 152 newborns enrolled from the Premaldo cohort were assessed.17 Briefly, inclusion criteria were mothers aged 18–45 years old, with no history of type 1 or 2 diabetes and none of the following maternal treatment prior to pregnancy (systemic or inhaled glucocorticoid therapy, hormonal treatment for adrenal or pituitary insufficiency and antihypertensive drugs). Infants with a birth weight between the 10th and 90th percentile for gestational age (GA) were subsequently enrolled. Three groups were defined, according to GA: group 1:<33 GW, VPT infants, n=46; group 2: 33–36 GW, moderate to late PT infants, n=67; and group 3: ≥37 GW, T infants, n=39. On day 3 of life, leftovers from 70 samples were assessed in group 1 (n=11), group 2 (n=28) and group 3 (n=31).
Study approval
Informed and written consent was obtained from both parents prior to inclusion. The study was conducted in accordance with the Declaration of Helsinki and approved by the ethics committee (Ile de France).
Blood samples
Venous blood samples were collected from umbilical cords at birth and from peripheral blood on day 3 into ethylenediaminetetracetic acid containing tubes. They were immediately transferred to the laboratory for centrifugation. Supernatants were stored at −80°C until analysis.
Steroid hormone analysis
Twelve steroids and precursors of adrenal pathways were assessed by a LC-MS/MS method: aldosterone (aldo), 18-hydroxycorticosterone (18-OH-B), corticosterone (B), 11-deoxycorticosterone (DOC), progesterone, 17-hydroxyprogesterone (17-OH-P), 11-deoxycortisol (S), cortisol (F), cortisone (E), 21-deoxycortisol (21DF), DHEAS and androstenedione (D4). The limits of quantification were 0.015 ng/mL for aldo, 0.050 ng/mL for DOC, 17-OH-P and B and 1.000 ng/mL for cortisol.18 The enzymatic activity of each biosynthesis step was estimated by the product-to-substrate ratio and referred to as enzymatic activity index (EAI).19
Statistics
All steroid concentrations and EAI are expressed as median and IQR. Non-parametric analysis of variance Kruskall-Wallis tests were used followed by Tukey post-tests for comparison between groups. Correlations between GA and hormone concentrations or EAI were assessed by Spearman correlation tests. All statistics were calculated using GraphPad Prism software (San Diego, California, USA). A p value of less than 0.05 was considered as statistically significant.
Results
Global immaturity of steroid biosynthesis at birth in VPT infants
Mineralocorticoid pathway
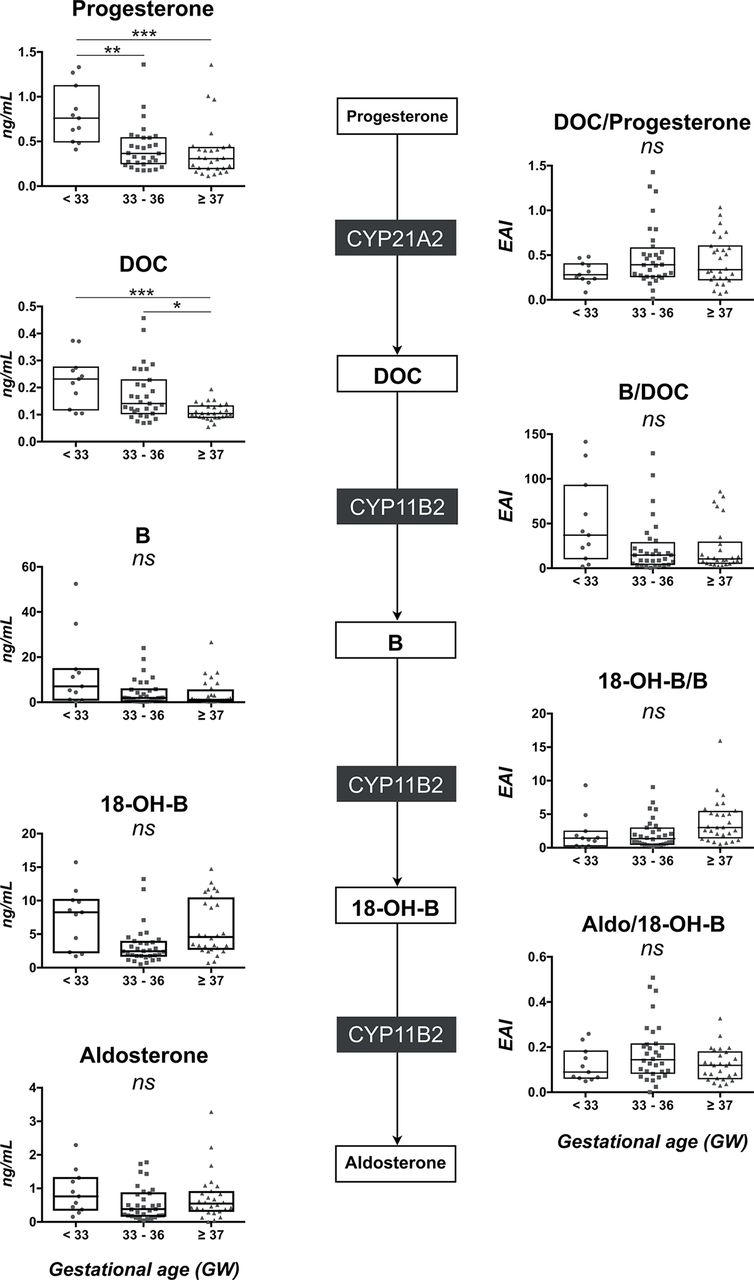
As expected, progesterone levels were above the upper limit of linearity (50 ng/mL) and DOC levels were similar in the three groups at birth. As compared with term neonates, a global defect was observed for B, 18-OH-B and aldosterone levels in VPT neonates (figure 1, left panels and see online supplementary table 1). Hormone levels were correlated with GA for B (r=0.22, p<0.02), 18-OH-B (r=0.32, p<0.001) and aldosterone (r=0.25, p<0.01). The EAI was calculated for each enzymatic reaction. The 11-hydroxylase activity of CYP11B2 (B/DOC) was lower in VPT (0.86 (0.62–1.27)) than in moderately PT (1.33 (0.83–2.21), p<0.05) and T neonates (1.26 (0.78–2.26), p<0.05). However, 18-hydroxylase (18-OH-B/B) and 18-oxidase (aldo/18-OH-B) activities of CYP11B2 were similar in the three groups (figure 1, right panels).
Supplemental material

The mineralocorticoid pathway in the umbilical cord blood. On the left panel, comparison of DOC, B, 18-OH-B and aldosterone concentrations (scatter plot with median and IQR) between the three groups of neonates (very preterm <33 GW, moderately preterm 33–36 GW and term neonates ≥37 GW). On the right panel, comparison of the product-to-substrate ratio as an index for each enzymatic step between the three groups of neonates. Statistical significance (Kruskal-Wallis and post hoc Tukey tests): ns, not significant; *p<0.05; **p<0.01; ***p<0.001; ****p<0.0001. 18-OH-B, 18-hydroxycorticosterone; B, corticosterone; DOC, 11-deoxycorticosterone; GW, gestational week.
Glucocorticoid pathway
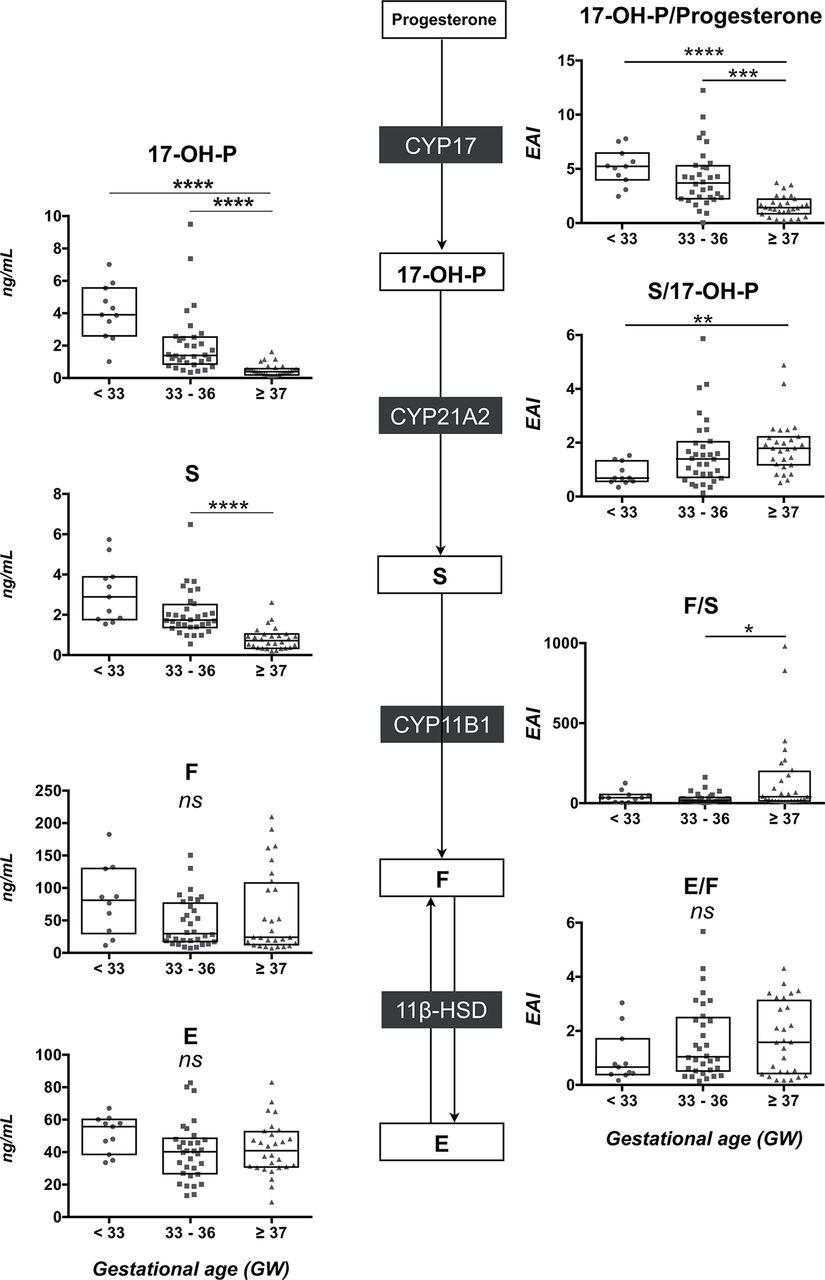
A global defect of glucocorticoid synthesis was also observed in VPT neonates. Cord blood concentrations of 17-OH-P, S, F and E were lower in PT and VPT compared with T neonates (figure 2, left panels and see online supplementary table 1). Hormone levels were strongly correlated with GA for 17-OH-P (r=0.35, p=0.0002), S (r=0.35, p=0.0002) and F (r=0.50, p<0.0001).

The glucocorticoid pathway in the umbilical cord blood. On the left panel, comparison of 17-OH-P, S, cortisol and E concentrations (scatter plot with median and IQR) between the three groups of neonates (very preterm <33 GW, moderately preterm 33–36 GW and term neonates ≥37 GW). On the right panel, comparison of the product-to-substrate ratio as an index for each enzymatic step between the three groups of neonates. Statistical significance (Kruskal-Wallis and post hoc Tukey tests); ns, not significant; *p<0.05; **p<0.01; ***p<0.001; ****p<0.0001. 17-OH-P, 17-hydroxyprogesterone; F, cortisol; E, cortisone; S, 11-deoxycortisol; GW, gestational week; .
The 21-hydroxylase activity (S/17-OH-P) was similar in the three groups (figure 2, right upper panel) and the 11-hydroxylase activity (F/S) was lower in VPT (5.5 (4.2–9.1), p<0.01) and PT (7.1 (4.0–10.1), p<0.01) than in T neonates (16.5 (7.1–20.2)) (figure 2, right lower panel). In addition, the E/F ratio evaluating the combined 11β-HSD1/HSD2 activities was higher in VPT (4.5 (2.7–7.1), p<0.01) and PT (5.1 (3.1–7.3), p<0.001) than in T neonates (2.6 (1.9–3.9)).
Increased steroid precursors in VPT infants at day 3
Mineralocorticoid pathway
As expected, plasma progesterone levels dramatically dropped in all groups from above 50 ng/mL to below 1 ng/mL (figure 3, left panels and see online supplementary table 1). However, progesterone and DOC concentrations remained significantly higher in VPT than in T neonates and were negatively correlated to GA (r=−0.45, p=0.03 and r=−0.42, p=0.003, respectively). In contrast, all steroid concentrations from B to aldosterone were similar in the three groups (figure 3, left panels and see online supplementary table 1). No relationship was observed between B, 18-OH-B or aldosterone levels and GA.

The mineralocorticoid pathway in peripheral blood on day 3. On the left panel, comparison of progesterone, DOC, B, 18-OH-B and aldosterone concentrations (scatter plot with median and IQR) between the three groups of neonates (very preterm <33 GW; moderately preterm 33–36 GW and term neonates ≥37 GW). On the right panel, comparison of the product-to-substrate ratio as an index for each enzymatic step between the three groups of neonates. Statistical significance (Kruskal-Wallis and post hoc Tukey tests): ns, not significant; *p<0.05; **p<0.01; ***p<0.001; ****p<0.0001. 18-OH-B, 18-hydroxycorticosterone; B, corticosterone; DOC, 11-deoxycorticosterone; GW, gestational week.
The EAI of 21-hydroxylase (DOC/progesterone), 11-hydroxylase (B/DOC), 18-hydroxylase (18-OH-B/B) and 18-oxidase (aldo/18-OH-B) were similar in the three groups (figure 3, right panels). Moreover, GA was neither correlated with 21-hydroxylase activity (DOC/progesterone: r=0.20, p=0.18) nor with 11-hydroxylase activity on day 3 (B/DOC: r=0.23, p=0.13).
Glucocorticoid pathway
As presented in figure 4 (left panels and see online supplementary table 1), 17-OH-P and S concentrations were higher in VPT and PT than in T neonates, whereas F and E levels were similar in the three groups (see online supplementary table 1). Moreover, strong negative relationships were observed between 17-OH-P or S levels and GA (r=−0.75, p<0.0001 and r=−0.76, p<0.0001, respectively), while no correlation was observed between F levels and GA (figure 5). The EAI of 21-hydroxylase (S/17-OH-P) was lower in VPT than in T neonates (figure 4, right middle panel). Nevertheless, this ratio was not related to GA (r=0.27, p=0.19) and 21DF levels remained undetectable in the three groups. Concerning the 11-hydroxylase activity, the F/S ratio was lower in PT (34.2 (15.7–50.4)) than in T neonates (40.5 (15.4–183.4), p<0.05) and strongly correlated with GA (r=0.58, p<0.0001) (figure 5). Finally, the E/F ratio was similar in the three groups (figure 4, right lower panel).
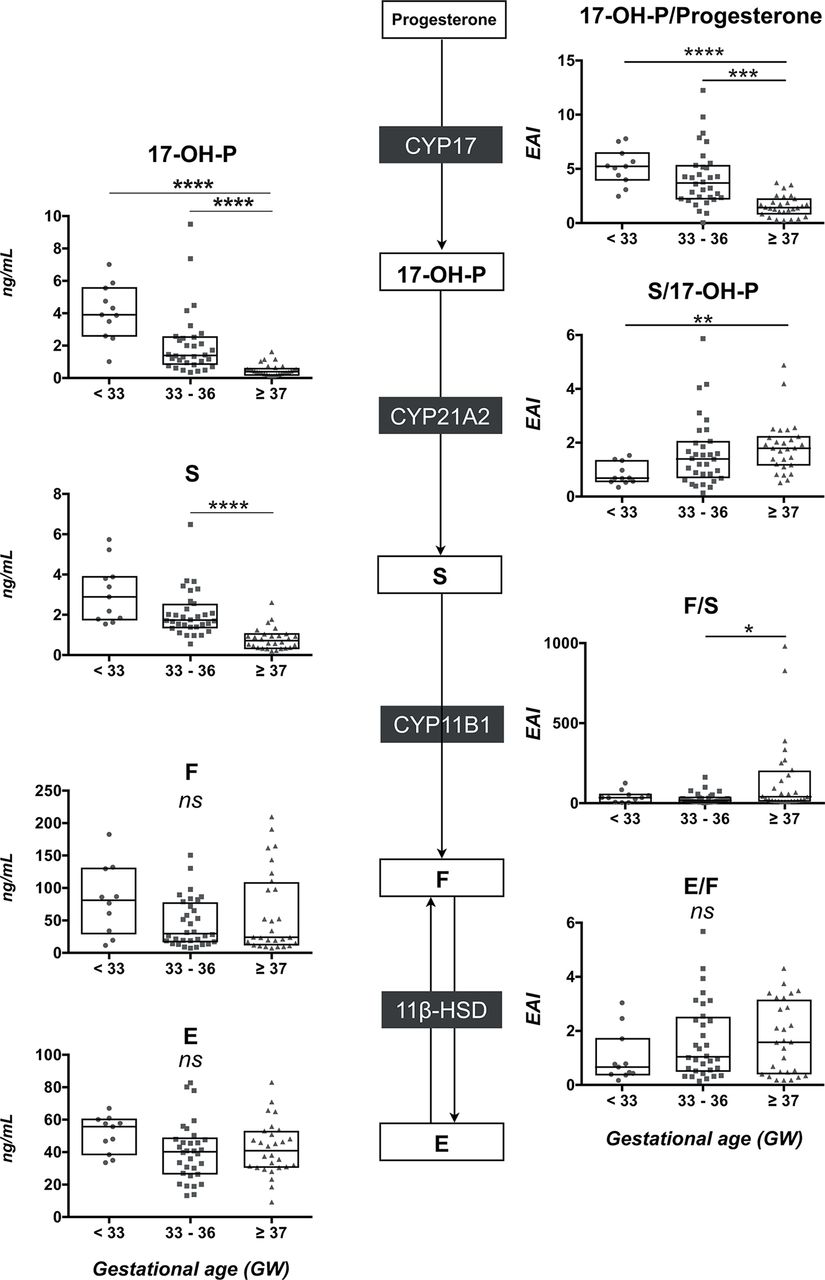
The glucocorticoid pathway in peripheral blood on day 3. On the left panel, comparison of 17-OH-P, S, F and E concentrations (scatter plot with median and IQR) between the three groups of neonates (very preterm <33 GW; moderately preterm 33–36 GW and term neonates ≥37 GW). On the right panel, comparison of the product-to-substrate ratio as an index for each enzymatic step between the three groups of neonates. Statistical significance (Kruskal-Wallis and post hoc Tukey tests): ns, not significant; *p<0.05, **p<0.01; ***p<0.001; ****p<0.0001. 17-OH-P, 17-hydroxyprogesterone; E, cortisone; F, cortisol; S, 11-deoxycortisol; GW, gestational week;

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Correlations between 11-deoxycortisol (S) and cortisol (F) concentrations in peripheral blood and gestational age on day 3. The product-to-substrate ratio (F/S) evaluating the 11-hydroxylase activity (CYP11B1) was positively correlated with the gestational age. Statistical significance (Spearman correlation tests): *p<0.05; **p<0.01; ***p<0.001; ****p<0.0001; ns, not significant.
Evolution of circulating androgens during the first 3 days of life
As reported in online supplementary table 1, androstenedione and DHEAS levels sharply decreased in T neonates between birth and day 3 while they rose in VPT neonates. At birth, cord blood levels of androstenedione and DHEAS were lower in VPT and PT than in T infants. Moreover, GA was strongly correlated with androstenedione (r=0.34, p=0.0002) and DHEAS (r=0.32, p=0.001) levels. In contrast, on day 3, androstenedione and DHEAS levels were higher in VPT than in T neonates, while androstenedione negatively correlated with GA (r=−0.38, p=0.01).
Discussion
In the present study, we establish for the first time the complete adrenal steroid profile using a highly sensitive and specific LC-MS/MS technology in a cohort of premature and term neonates. We provide evidence that prematurity is associated with alterations of corticosteroid biosynthesis pathways during the first 3 days of life. The profiling of adrenal steroids in cord blood shows large amounts of steroid precursors, progesterone and 17-OH-P. Despite this excess of substrates, VPT infants exhibit a global defect in both aldosterone and cortisol, strongly correlated with GA. A similar relationship between umbilical cord cortisol and GA has been previously reported in very low birth weight infants.8 On day 3, steroids from placenta sharply drop. Both aldosterone and cortisol concentrations normalise in VPT neonates with, however, persistence of high circulating levels of steroid precursors.
Several factors may be implicated in this global hormonal defect at birth in VPT infants. First, the placenta is a source of large quantities of progesterone and corticotrophin-releasing hormone (CRH). Placental CRH increases exponentially in both maternal and fetal blood as the gestation progresses towards term. During the third trimester, CRH stimulates DHEAS and cortisol production by fetal adrenal glands. At birth, placental supply of CRH and progesterone to the fetus is suddenly discontinued. Thus, the hypothalamic–pituitary–adrenal axis (HPA) may be transiently refractory and unable to stimulate steroid production in VPT infants.20 Additionally, the fetal adrenal cortex lacks certain enzymes essential for steroidogenesis such as 3β-hydroxysteroid dehydrogenase (3β-HSD) until 24 GW.21 22 Cortisol synthesis by fetal adrenals uses high levels of placental progesterone as a precursor to converse to 17-OH-P, bypassing the need for 3β-HSD.23 Therefore, the brutal fall of this placental substrate may contribute to the global yet partial steroid deficiency at birth.
The other main regulator of fetal circulating cortisol (F) levels is the placental 11β-hydroxysteroid dehydrogenase-2 (11βHSD2) that converts both F and B to their inactive 11-reduced metabolites, cortisone (E) and 11-dehydrocorticosterone (A), respectively.24 As assayed in cord blood, 11βHSD2 activity was higher in PT infants while it became similar on day 3 (figures 2 and 4). Similarly, Kajantie et al have reported a negative relationship between GA and placental 11βHSD2 activity.25 Thereby, this massive placental 11βHSD2 activity partly inactivates both cortisol and corticosterone towards their 11-keto derivatives and thus may contribute to dampen cortisol and aldosterone levels in cord blood. Normalisation of both hormones few days after birth also supports this hypothesis.
Another alternative explanation is the impact of antenatal corticosteroid exposure on the neonatal HPA axis activity. Ninety-three per cent of VPT and 78% of PT infants received antenatal glucocorticosteroids to promote lung maturation of the newborn. Indeed, the degree of suppression of the HPA axis is directly proportional to the dose of antenatal glucocorticosteroids and inversely proportional to the delay between the last administered dose and the delivery.8 26 Previous studies have shown that antenatal glucocorticosteroids reduce aldosterone, cortisol or 17-OH-P levels in children born 1–3 days after the last administered dose.27–30 Therefore, we could not exclude that such a negative impact of antenatal glucocorticosteroid administration may contribute for the reduced aldo and F levels in VPT.
On day 3, progesterone and 17-OH-P levels markedly fall in peripheral blood owing to placenta withdrawn. Nevertheless, compared with T neonates, higher levels of steroid precursors (progesterone, DOC, 17-OH-P, S and androstenedione) persist in VPT neonates (figure 3 & figure 4 and see online supplementary table 1). This is also supported by the strong and negative relationship between their blood concentrations and GA (figure 5).
A failure of an enzymatic step of adrenal biosynthesis might account for this larger excess of precursor’s steroids in VPT infants. Some studies have reported a decreased 21-hydroxylase activity in PT infants.31 However, the S/17-OH-P ratio is similar in the three groups at birth. On day 3, the S/17-OH-P ratio is lower in VPT than in T neonates but without any correlation with GA. Furthermore, a CYP21A2 deficit is unlikely because, first, 21-DF levels remained undetectable in all neonates and second, the expression of the enzyme appears early during gestation in all three zones of the fetal adrenals.32 33
Concerning the next step of biosynthesis pathway, the diminished F/S ratio is consistent with a decreased activity of 11β-hydroxylase in VPT infants (figures 2, 4 and 5). Previous investigators have already reported a similar decreased F/S ratio in the blood of premature infants7 10 12 and from the urinary steroid metabolome of PT or T neonates.11 This potential decrease in 11β-hydroxylase activity is also supported by the modest expression of CYP11B1 transcripts from 22 to 29 GW in human adrenals.33 Finally, despite higher levels of steroid precursors, a marked increase in cortisol and aldosterone blood levels was observed in VPT neonates between the birth and day 3 (figure 5), as also previously reported in healthy or sick PT neonates.7 9 34 35 In 50 neonates of less than 30 GW, Bourchier reported on D1 high plasma aldosterone levels inversely related to illness severity and 24 hours sodium intake suggesting a distal renal tubular unresponsiveness during the first days of life of VPT.36 In contrast, we previously reported in our cohort a strong relationship between urinary aldosterone and urinary Na/K ratio that demonstrated the conserved renal aldosterone sensitivity in VPT neonates.17
Several hypotheses can be put forward. First, VPT neonates have reduced biotransformation capacity of their liver.37 Indeed, Homma et al suggested a lowest activity of metabolising enzymes such as hepatic 5-reductase that could lead to compromised conversion to tetra-hydro-derivatives in VPT infants.38 This hypothesis is currently being investigated in urinary samples of neonates from the Premaldo cohort. However, at birth, the 11βHSD2 activity is suddenly collapsed with placenta withdrawn. As this 11βHSD2 activity was higher in VPT neonates before delivery, its disappearance might boost the levels of corticosterone and cortisol in VPT compared with T neonates.
Our study has obviously some limitations. First, it is well established that cord blood samples contain mixed analytes from maternal, placental and fetal origins, which are indistinguishable. Furthermore, samples were collected at delivery and measurements may not represent analyte levels earlier in pregnancy. Second, the low number of patient samples on day 3 is explained by the fact that we used leftovers. However, these 11 neonates were similar to the whole group VPT on D0 in term of GA and steroids profiles. Additional studies on a larger series of VPT neonates are needed to confirm our findings. Finally, this study underlines the limits in interpreting the product-to-substrate ratio as reflecting enzyme activity: a steroid being both substrate and product of different enzymatic steps.
In summary, VPT infants exhibit at birth a global defect of adrenal steroid pathways resulting in diminished levels of aldosterone, cortisol and androstenedione. On day 3, these levels return to normal values while circulating steroid precursors accumulate in VPT neonates. Furthermore, this study provides additional support for a partial defect in 11-hydroxylase activity at birth. Since immaturity is associated with adverse outcomes in premature infants, it is likely amendable by appropriate pharmacological options that remain to be evaluated.
Acknowledgments
ST was recipient of a fellowship from the French Endocrine Society. This work has been supported by fundings from Institut National de la Santé et de la Recherche Médicale (INSERM), Université Paris Sud, Assistance Publique de Paris. We also acknowledged the funding from a Cardiovasculaire-Obésité-Rein-Diabète-Domaine d’Intérêt Majeur (CORDDIM, Région Ile-de-France) grant that partly supported the acquisition of the Waters Xevo TQS triple-quadrupole mass spectrometer.
References
Footnotes
ML and EP contributed equally.
Contributors All authors contributed to the design of the study. ST and EP performed the analyses. All authors analysed and interpreted the data. ST, ML and EP wrote a first draft of the manuscript, and all coauthors critically evaluated and suggested revisions to the manuscript and approved the final submission.
Competing interests None declared.
Patient consent Obtained.
Ethics approval CCP Comitè de protection des personnes, Ile de France.
Provenance and peer review Not commissioned; externally peer reviewed.
Linked Articles
- Fantoms