Article Text

Abstract
Objective To determine if preterm birth is associated with adaptation of the hypothalamic–pituitary–adrenal (HPA) axis and whether HPA axis programming relates to the degree of prematurity (defined as extremely preterm birth at <28 weeks or very preterm birth at 28–32 weeks gestation).
Design This study reports findings from a prospective birth cohort. Saliva cortisol concentrations were measured prevaccination and postvaccination, and in the morning and evening, at 4 months chronological age.
Setting Infants born at a single Scottish hospital.
Participants 45 term-born, 42 very preterm and 16 extremely preterm infants.
Outcomes Cortisol stress response to vaccination (postvaccination minus prevaccination cortisol concentrations), diurnal slope (log-transformed morning minus log-transformed evening cortisol values) and mean log-transformed daily cortisol.
Results Compared with infants born at term, infants born extremely preterm had a blunted cortisol response to vaccination (5.8 nmol/L vs 13.1 nmol/L, difference in means: −7.3 nmol/L, 95% CI −14.0 to −0.6) and a flattened diurnal slope (difference in geometric means: −72.9%, 95% CI −87.1 to −42.8). In contrast, the cortisol response to vaccination (difference in means −2.7 nmol/L, 95% CI −7.4 to 2.0) and diurnal slope at 4 months (difference in geometric means: −33.6%, 95% CI −62.0 to 16.0) did not differ significantly in infants born very preterm compared with infants born at term.
Conclusions Infants born extremely preterm have blunted cortisol reactivity and a flattened diurnal slope. These patterns of HPA axis regulation are commonly seen after childhood adversity and could contribute to later metabolic and neurodevelopmental phenotypes observed in this population.
- neonatology
- endocrinology
- infant development
This is an open access article distributed in accordance with the Creative Commons Attribution 4.0 Unported (CC BY 4.0) license, which permits others to copy, redistribute, remix, transform and build upon this work for any purpose, provided the original work is properly cited, a link to the licence is given, and indication of whether changes were made. See: https://creativecommons.org/licenses/by/4.0/.
Statistics from Altmetric.com
What is already known on this topic?
Preterm birth is associated with adverse metabolic and neurodevelopmental phenotypes across the life course.
Hypothalamic–pituitary–adrenal axis (HPA) dysregulation has been observed following multiple types of prenatal and childhood adversity and is a potential biological mediator of later pathology.
What this study adds?
At 4 months postnatal age extremely preterm birth is associated with a blunted cortisol response to vaccination and a flattened diurnal rhythm.
Future studies investigating whether HPA axis adaptations after preterm birth continue across the life course and relate to adverse neurodevelopmental and metabolic phenotypes appear warranted.
Introduction
Preterm birth (<37 weeks gestation) is associated with an increased risk of adverse health across the life course, including neurodevelopmental impairment,1 2 inattention, mood disorders and psychosis,3 metabolic disorders,4 and cardiovascular disease.5 However, the biological mechanisms linking preterm birth with future morbidity remain largely unknown.
The hypothalamic–pituitary–adrenal (HPA) axis, the primary regulator of human endogenous cortisol secretion, is a potential biological mediator between early life adversity and later adverse neurodevelopmental and cardiometabolic phenotypes. Its candidacy is supported by a growing body of preclinical and human observation research that demonstrates a complex interplay between early life adversity, adaptations in cortisol regulation and health across the life course.6 Epidemiological studies commonly use saliva cortisol, which has been validated as a non-invasive marker of serum cortisol concentrations through many studies.7
Previous studies of infant HPA axis regulation after preterm birth have typically focused on stress reactivity. There is emerging evidence that infants born preterm may have a blunted cortisol response to physiological and psychological stressors8 9 and that this potentially relates to the degree of prematurity10 and sex.11
To date, only one study has assessed basal cortisol of term and preterm infants across infancy. Infants born between 28 and 32 weeks (very preterm) and <28 weeks (extremely preterm) had lower cortisol concentrations than term infants at 3 months corrected age, with extremely preterm infants having higher levels at 8 months.12 Assessments were made using single saliva collections at each timepoint, which is unlikely to be representative of cortisol output across the day. Additionally, single measurements do not allow for characterisation of diurnal rhythmicity. In adults cortisol concentrations typically peak in the morning before falling across the day, and the extent of this decline can be described as the diurnal slope.
This study presents data from a prospective birth cohort conducted with the primary aim of investigating whether preterm birth predisposes infants to differences in stress-induced and diurnal cortisol release in infancy. We hypothesised preterm birth would predispose infants to a blunted stress response to vaccination and flatter decline in cortisol across the day.13 14 Given the complications of preterm birth are gestation-specific,15 with adverse effects more likely for infants born at younger gestations, we hypothesised HPA axis programming effects would be most pronounced in infants born extremely preterm. Additionally, we sought to assess if HPA axis programming after preterm birth was sex-specific.
Methods
Participants
The ‘Stress Response Systems in Mothers and Preterm Infants’ study recruited infants born at the Royal Infirmary of Edinburgh, UK, between March 2018 and August 2019. All parents gave written informed consent. Exclusion criteria were regular maternal steroid use in pregnancy and congenital chromosomal or structural abnormalities in infants.
At 4 months chronological age infants had four saliva samples collected at the time of routine scheduled vaccinations against diphtheria, tetanus, pertussis, polio,haemophilus influenzae type b and hepatitis B (DTaP/IPV/Hib/HepB), meningitis B and pneumococcus (13 serotypes). Vaccinations were administered as three intramuscular injections at participants’ general practitioner surgeries, as per routine clinical care. Appointments were attended by a member of the research team, who collected samples immediately before and 20 min after vaccinations, assessing cortisol reactivity. Morning (07:30–09:30) and evening (19:30–21:30) samples were collected by participants’ parents in their own homes, assessing diurnal cortisol regulation. Parents were asked to collect these samples on a single day in the week preceding the vaccination and to refrigerate samples before transferring them to the research team at the vaccination appointment. Saliva was collected using Salimetrics SalivaBio Infant Swabs, stored at −80°C and analysed by liquid chromatography tandem mass spectrometry (LC-MS/MS) at Edinburgh University Clinical Research Facility Mass Spectrometry Core (online supplemental appendix 1).
Supplemental material
Statistical analysis
Analyses were performed using IBM SPSS Statistics V.25. Continuous data are summarised as mean±SD if normally distributed and median (IQR) if skewed. One participant concurrently using topical steroids was excluded from analysis. One morning cortisol sample was also excluded (as it had a supraphysiological cortisol concentration at >50 nmol/L). Distributions of cortisol concentrations were assessed for normality using histograms. Cortisol concentrations for the assessment of diurnal cortisol regulation were positively skewed and log10-transformed for further analysis. Prevaccine cortisol concentrations were positively skewed. However, vaccine cortisol concentrations were not transformed in the presented analysis as postvaccine cortisol concentrations and subsequent regression model residuals showed only slight asymmetry. Cortisol distributions across all timepoints are presented through violin plots in figure 1.

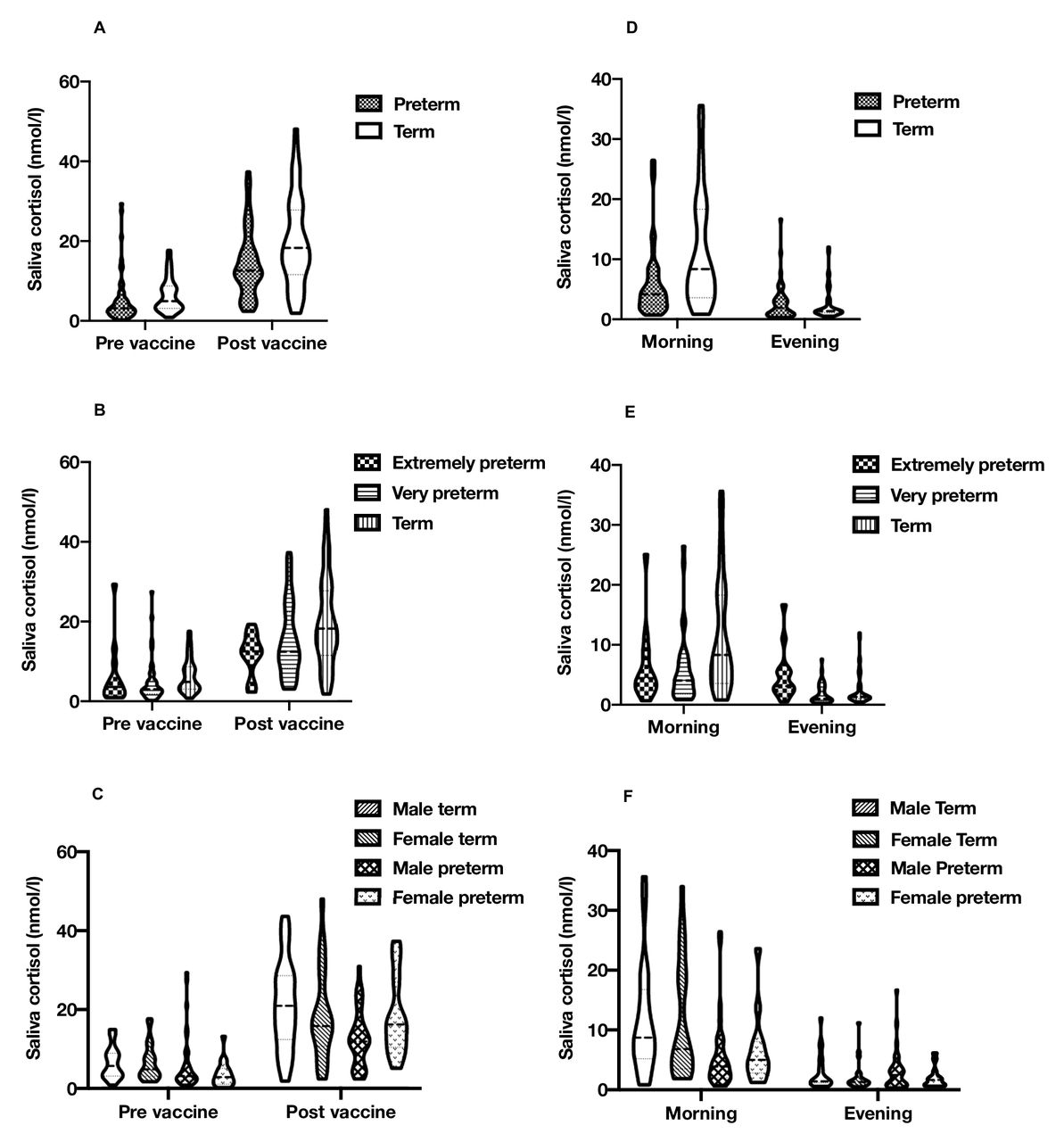{kind=link}
Violin plots of saliva cortisol levels. Range, median and quartiles for (A) term/preterm vaccine response (n=45 vs 56); (B) term/extremely/very preterm vaccine response (n=45 vs 42 vs 14); (C) sex/birth group vaccine response (n=21 vs 24 vs 38 vs 18); (D) term/preterm diurnal cortisol (n=42 vs 48); (E) term/extremely/very preterm diurnal cortisol (n=42 vs 34 vs 14); and (F) sex/birth group diurnal cortisol (n=20 vs 22 vs 35 vs 13). The figure was produced by DQS using GraphPad Prism V.8.4.3.
Comparisons of HPA axis regulation between term and preterm infants (defined as a single group) and preterm subgroups (defined as extremely preterm birth at <28 weeks or very preterm birth at 28–32 weeks gestation) were conducted using linear mixed models. Cortisol reactivity in response to vaccination and diurnal cortisol patterns (average cortisol across the day and diurnal slope) were tested separately. In each model, time (defined as prevaccination=0 and postvaccination=1, or evening=0 and morning=1) was specified as a repeated measure. Preterm birth’s influence on cortisol reactivity and diurnal cortisol decline across the day was tested by the time*preterm/term group interaction. To test if HPA axis programming is limited, or differs by degree of prematurity, the time*extremely preterm/very preterm/term interaction was assessed. To assess if associations between preterm birth and HPA axis regulation differed by sex, the time*sex*preterm/term interaction was tested. This process was also undertaken for assessment of mean diurnal cortisol levels, using morning and evening cortisol concentrations in models that did not include an interaction with time.
All analyses were conducted with models specifying a diagonal covariance structure (assuming no association between and unequal variance of the repeated measures). This is because the variance of cortisol concentrations at the different timepoints differed, and models incorporating a correlation in cortisol concentrations between participant samples did not improve the model fit assessed through comparison of −2 log likelihood ratios.
Sensitivity analyses were conducted assessing the potential effect of comparing infants at chronological compared with corrected ages. In this study, where saliva was collected at 4 months chronological age, the corrected age of saliva samples collected in the morning and evening (Pearson’s correlation coefficient, r=0.908) and around the vaccination (r=0.876) was strongly correlated with birth gestation. As corrected age at saliva sampling could not directly be adjusted for in models without introducing multicollinearity, the potential influence of corrected age at sampling on cortisol concentration was assessed through univariate analysis in term and preterm groups separately.
Effect sizes are reported as mean differences in cortisol concentrations for normally distributed data and percentage differences of geometric means for skewed data, with 95% CI. P<0.05 was considered statistically significant.
Results
Demographics
Infant and sampling demographics are presented in table 1. As sample collection was timed according to when infants received their third set of routine vaccinations (and the vaccine schedule is scheduled according to postnatal age), corrected age at sampling differed between birth groups. Cortisol reactivity to vaccination was assessed at a median of 5.6 weeks corrected age (IQR 3.7–11.8) in extremely preterm infants and 9.0 weeks (IQR 8.1–11.8) in very preterm infants.
Study demographics
Cortisol response around vaccination
Salivary cortisol increased in response to vaccination in all groups (p<0.001) (table 2). Compared with infants born at term, no difference in cortisol reactivity was observed in the combined preterm group (−3.9 nmol/L, p=0.09) (figure 1A) or in very preterm infants (−2.7 nmol/L, p=0.26) (figure 1B). In contrast, cortisol reactivity in response to vaccination was blunted in extremely preterm infants (−7.3 nmol/L, p=0.03). There was also an interaction between birth group and sex (p=0.02). Male preterm infants had a blunted cortisol response compared with male term infants (−7.7 nmol/L, p=0.009), but female infants did not show this pattern of response (2.5 nmol/L, p=0.47) (figure 1C).
Cortisol reactivity to vaccination
Diurnal cortisol
At 4 months chronological age, cortisol concentrations reduced across the day, with higher cortisol in the morning than evening (p<0.001) (table 3). Compared with infants born at term, preterm infants had a reduced mean cortisol across the day (−26.8%, p=0.02) and a flattened diurnal slope (−48.8%, p=0.01) (figure 1D). In comparison with term infants, extremely preterm infants had a flattening of diurnal slope (−72.9%, p<0.001) but no difference in mean concentrations across the day (13.3%, p=0.52), with very preterm infants having lower mean levels (−36.6%, p=0.002), but not a significant flattening of the diurnal slope (−33.6%, p=0.15) (figure 1E). In a post-hoc analysis conducted to interrogate these differing patterns of diurnal cortisol observed between the preterm groups, extremely preterm (p=0.04) and very preterm (p=0.002) infants both had lower morning cortisol concentrations than term infants, while only infants born extremely preterm (p=0.006) had higher evening cortisol concentrations. Sex did not interact with birth group and when testing the associations for diurnal slope (p=0.78) or mean levels across the day (p=0.62) (figure 1F).
Diurnal cortisol
Sensitivity analyses
Mean diurnal cortisol concentrations were negatively associated with corrected age at sampling in term (p=0.02) but not in preterm (p=0.73) infants. Cortisol diurnal slope and reactivity to vaccination were not associated with corrected age at sampling in either the term or preterm group (online supplemental table 1).
Discussion
This study has two main findings. First, extremely preterm birth is associated with altered HPA axis regulation, including both blunted stress response and flattened diurnal slope in infancy. Second, infant sex was related to HPA axis programming of cortisol reactivity but not diurnal cortisol concentrations.
The ‘developmental origins of health and disease’ (DOHaD) hypothesis postulates that environmental exposures in early life, during periods critical for development, can influence health across the life course.16 Interest in this concept has expanded since it was shown that reduced fetal size, a surrogate for an adverse in utero environment, is associated with insulin resistance17 and cardiovascular disease in adulthood.18
At 4 months chronological age extremely preterm infants had a reduced cortisol response to vaccination and a reduced decline in cortisol across the day. While our primary hypothesis was that infants born ≤32 weeks gestation would show adaptation of the HPA axis, an increased propensity for HPA axis programming in infants born extremely preterm appears physiologically plausible, when viewed through a DOHaD lens, as this group face an earlier disruption to environment and greater morbidity in the newborn period. Additionally, evidence from previous studies provides precedent that HPA axis programming may be gestation-specific.10 12
This study adds to evidence that early exposure to an ex utero environment contributes to blunted cortisol reactivity across life.8–10 19 20 Blunting of cortisol reactivity in male compared with female preterm infants is consistent with a previous observation of cortisol reactivity of preterm infants in response to vaccination11 and adds to evidence of vulnerability of the male axis after preterm birth.21–23
This study is the first to address whether preterm birth influences diurnal slope in infancy. However, available data from studies in childhood are indicative of a flattening of the HPA axis after preterm birth, including two reports of reduced morning cortisol24 25 and one of increased evening cortisol concentrations.23 Furthermore, a flatter diurnal cortisol slope and blunted cortisol reactivity to stressors are patterns of HPA axis regulation frequently observed after childhood adversity.13 14 26
The altered patterns of saliva cortisol observed in this study are of clinical interest as HPA axis dysregulation is potentially causative of cardiometabolic27 and neurobehavioural pathology.28 Importantly evidence is emerging that flattening of the diurnal slope after early life stress is reversible, and so this may be a target for future intervention.29
Multiple perinatal exposures hold the potential to contribute to programming of the HPA axis in a preterm population. These include intrauterine and extrauterine nutritional deficiencies and growth restriction,30 chorioamnionitis and postnatal septic episodes,31 and noxious environmental stimuli.21 32
Additionally, perinatal hormonal exposures could change the way that cortisol secretion is regulated at the level of the hypothalamus or pituitary glands across infancy, or alter the development of the adrenal cortex. Preterm infants are commonly exposed to exogenous steroids and higher endogenous cortisol concentrations than would be expected at a comparative gestation in utero,33 and a blunted stress response and flattened diurnal cortisol could reflect adaptations of the HPA axis in response to these exposures.
Preterm infants also have reduced exposure to hormones typically secreted during the third trimester by the placenta, such as reduced corticotropin-releasing hormone, oestrogen and kisspeptin exposure. Reduced exposure to these hormones likely changes the developmental trajectory of the adrenal gland,34 35 an organ that undergoes considerable remodelling after birth with reduction in the androgen-producing fetal zone and maturation of the cortisol-producing zona fasciculata.
This study’s primary strength is that it combines assessment of both cortisol stress response and diurnal rhythm, providing an indepth characterisation of the preterm HPA axis in infancy. Additionally cortisol concentrations were quantified using LC-MS/MS using a long 16 min chromatographic analysis, such that temporal separation confidently excludes contribution by cortisone and corticosterone and its 11-dehydrocorticosterone, while distinct mass differences of cortisol (~362 Da) and androgens (~290 Da) allow quantification without interference by other adrenal steroids.
A potential limitation is that sampling was scheduled according to chronological age and so the corrected ages of participants in the extremely preterm group were younger than those at sampling of term group. There has been disagreement around whether adrenal function relates to postmenstrual age or time after birth, particularly around the involution of the fetal zone and continued production of fetal zone steroids.34 36 However, it has previously been demonstrated that both preterm and term-born infants typically have a higher morning than evening cortisol from 1 month corrected age, suggesting that corrected age may be more important for diurnal cortisol rhythms.37 38 Thus the younger gestational age of sampling in the cohort may have contributed to the flatter diurnal cortisol rhythms observed in extremely preterm compared with term-born infants. In contrast, it is unlikely that assessment at a younger corrected age contributed to the reduced cortisol response to vaccination observed in extremely preterm infants compared with term infants, as previous longitudinal studies of stress reactivity in infancy have demonstrated reductions, rather than increases, in cortisol reactivity with progressing age.33 39
Another limitation is that cortisol concentrations before the vaccination were higher than morning cortisol concentrations in the preterm groups. This suggests that attending the vaccination appointment may itself be a mild stressor. Additionally, the sample sizes of the extremely preterm group and female infants born preterm were relatively small, reducing the precision of effect size estimates for these groups.
Preterm birth has also been associated with adaptations in adrenal androgen secretion from the postnatal period into young adulthood,40 41 and these adaptations in turn hold further potential to influence neurodevelopmental and metabolic phenotypes. Future studies incorporating the measurement of both cortisol and adrenal androgens would enable a more complete understanding of adrenal function after preterm birth.
Conclusion
Extremely preterm birth disrupts the normative development of the HPA axis in infancy, with patterns of cortisol secretion resembling those seen after childhood adversity. Future research needs include testing how HPA axis adaptation relates to adverse neurodevelopmental and metabolic phenotypes seen after preterm birth and assessing whether psychosocial interventions can ‘reprogramme’ the preterm HPA axis.
Ethics statements
Patient consent for publication
Ethics approval
This study involves human participants and was approved by South East Scotland 01 Regional Ethics Committee (18/SS/0006). Participants gave informed consent to participate in the study before taking part.
Acknowledgments
We are grateful to the families who consented to participate in the study.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Twitter @EvalotteM
Contributors DQS conceptualised and designed the study, collected the data, carried out the initial analyses, drafted the initial manuscript, and reviewed and revised the manuscript. JPB conceptualised and designed the study and revised the article critically for important intellectual content. GS, GL and GSB acquired data and revised the article critically for important intellectual content. CO advised around statistical analysis and revised the article critically for important intellectual content. NZMH provided the analysis of saliva samples and revised the article critically for important intellectual content. NN, ET and EM contributed to the interpretation of data for the work and revised the article critically for important intellectual content. RMR conceptualised and designed the study, drafted the initial manuscript, and reviewed and revised the manuscript. All authors approved the final manuscript as submitted and agree to be accountable for all aspects of the work. RMR is guarantor.
Funding The work was funded by Theirworld (www.theirworld.org) and was undertaken at the MRC Centre for Reproductive Health, which is funded by MRC Centre Grant (MRC G1002033). RMR acknowledges the support of the British Heart Foundation (RE/18/5/34216). EM acknowledges Perth Children’s Hospital Foundation for supporting the professorial position.
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.